Cell Wall
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Bacteria
The bacterial cell wall is an extremely important structure, being essential for the maintenance of the shape and integrity of the bacterial cell. It is also chemically unlike any structure present in eukaryotic cells and is therefore an obvious target for antibiotics that can attack and kill bacteria without harm to the host.
CELL WALL
The bacterial cell wall is an extremely important structure, being essential for the maintenance of the shape and integrity of the bacterial cell. It is also chemically unlike any structure present in eukaryotic cells and is therefore an obvious target for antibiotics that can attack and kill bacteria without harm to the host.
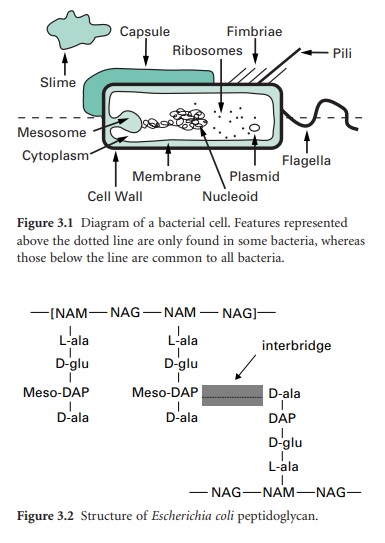
The primary function of the cell wall is to provide a strong, rigid structural component that can withstand the osmotic pressures caused by high chemical concentrations of inorganic ions in the cell. Most bacterial cell walls have in common a unique structural component called peptidoglycan (also called murein or glycopeptide); exceptions include the mycoplasmas, extreme halophiles and the archaea. Peptidoglycan is a large macromolecule containing glycan (polysaccharide) chains that are crosslinked by short peptide bridges. The glycan chain acts as a backbone to peptidoglycan, and is composed of alternating residues of N-acetyl muramic acid (NAM) and N-acetyl glucosamine (NAG). To each molecule of NAM is attached a tetrapeptide consisting of the amino acids L-alanine, D-alanine, D-glutamic acid and either lysine or diaminopimelic acid (DAP). This glycan tetrapeptide repeat unit is crosslinked to adjacent glycan chains, either through a direct peptide linkage or a peptide inter-bridge (Figure 3.2). The types and numbers of crosslinking amino acids vary from organism to organism. Other unusual features of the cell wall that provide potential antimicrobial targets are DAP and the presence of two amino acids that have the D-configuration.
Bacteria can be divided into two large groups, Gram-positive and Gram-negative, on the basis of a differential staining technique called the Gram stain. Essentially, the Gram stain consists of treating a film of bacteria dried on a microscope slide with a solution of crystal violet, followed by a solution of iodine; these are then washed with an alcohol solution. In Gram negative organisms the cells lose the crystal violet–iodine complex and are rendered colourless, whereas Gram-positive cells retain the dye. Regardless, both cell types are counter-stained with a different coloured dye, e.g. carbolfuchsin, which is red.
Hence, under the light microscope Gram-negative cells appear red while Gram-positive cells are purple. These marked differences in response reflect differences in cell wall structure. The Gram-positive cell wall consists primarily of a single type of molecule whereas the Gram-negative cell wall is a multi-layered structure and quite complex.
The cell walls of Gram-positive bacteria are quite thick (20–80 nm) and consist of between 60% and 80% peptidoglycan, which is extensively crosslinked in three dimensions to form a thick polymeric mesh (Figure 3.3). Gram-positive walls frequently contain acidic polysaccharides called teichoic acids; these are either ribitol phosphate or glycerol phosphate molecules that are connected by phosphodiester bridges. Because they are negatively charged, teichoic acids are partially responsible for the negative charge of the cell surface as a whole.
Their function may be to effect passage of metal cations through the cell wall. In some Gram-positive bacteria glycerol–teichoic acids are bound to membrane lipids and are termed lipoteichoic acids. During an infection, lipoteichoic acid molecules released by killed bacteria trigger an inflammatory response. Cell wall proteins, if present, are generally found on the outer surface of the peptidoglycan.
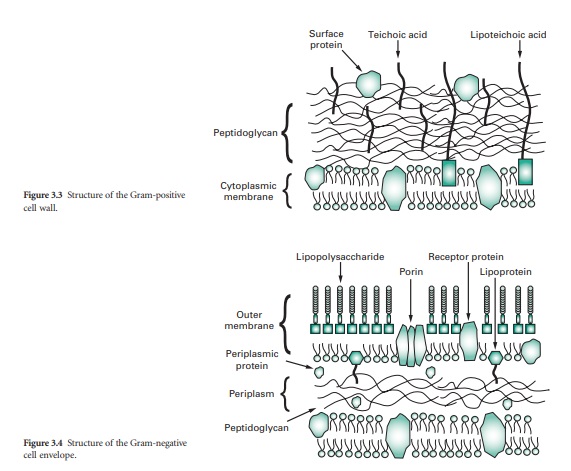
The wall, or more correctly, envelope of Gram-negative cells is a far more complicated structure (Figure 3.4). Although it contains less peptidoglycan (10–20% of wall), a second membrane structure is found outside the peptidoglycan layer. This outer membrane is asymmetrical, composed of proteins, lipoproteins, phospholipids and a component unique to Gram-negative bacteria, lipopolysaccharide (LPS). Essentially, the outer membrane is attached to the peptidoglycan by a lipoprotein, one end of which is covalently attached to peptidoglycan and the other end is embedded in the outer membrane. The outer membrane is not a phospholipid bilayer although it does contain phospholipids in the inner leaf, and its outer layer is composed of LPS, a polysaccharide–lipid molecule. Proteins are also found in the outer membrane, some of which form trimers that traverse the whole membrane and in so doing form waterfilled channels or porins through which small molecules can pass. Other proteins are found at either the inner or outer face of the membrane.
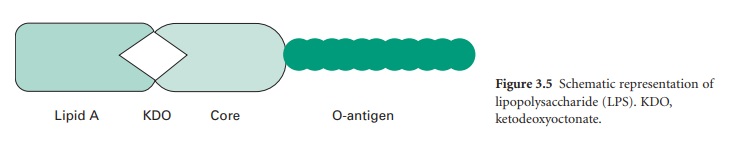
The LPS (Figure 3.5) is an important molecule because it determines the antigenicity of the Gram-negative cell and it is extremely toxic to animal cells. The molecule consists of three regions, namely lipid A, core polysaccharide and Ospecific polysaccharide. The lipid A portion is composed of a disaccharide of glucosamine phosphate bound to fatty acids and forms the outer leaflet of the membrane. It is the lipid A component that is responsible for the toxic and pyrogenic properties of Gram-negative bacteria. Lipid A is linked to the core polysaccharide by the unique molecule ketodeoxyoctonate (KDO), and at the other end of the core is the Opolysaccharide (Oantigen), which usually contains sixcarbon sugars as well as one or more unusual deoxy sugars such as abequose.
Although the outer membrane is relatively permeable to small molecules, it is not permeable to enzymes or large molecules. Indeed, one of the major functions of the outer membrane may be to keep certain enzymes that are present outside the cytoplasmic membrane from diffusing away from the cell. Moreover, the outer membrane is not readily penetrated by hydrophobic compounds and is, therefore, resistant to dissolution by detergents.
The region between the outer surface of the cytoplasmic membrane and the inner surface of the outer membrane is called the periplasm. This occupies a distance of about 12–15 nm, is gel like in consistency and, in addition to the peptidoglycan, contains sugars and an abundance of proteins including hydrolytic enzymes and transport proteins. Table 3.2 summarizes the major differences in wall composition between Gram-positive and Gram-negative cells.
Related Topics
