Cellular Reserve Materials
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Structure and Function of Bacterial Cells
It has been duly observed that there exist a good number of ‘reserve materials’ strategically located in the prokaryotic cells and are invariably known as the granular cytoplasmic inclusions.
Cellular Reserve Materials
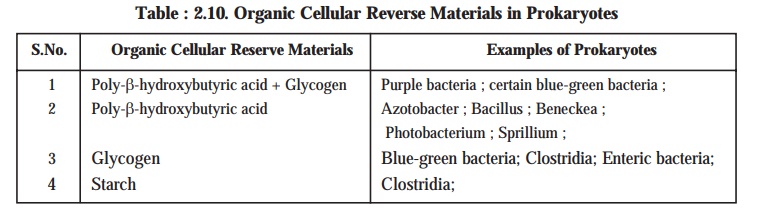
It has been duly observed that there exist a good number of ‘reserve materials’ strategically located in the prokaryotic cells and are invariably known as the granular cytoplasmic inclusions. The three most vital and important organic cellular reserve materials present in the prokaryotes are namely : (a) poly-β-hydroxybutyric acid; (b) glycogen; and (c) starch (see Table : 2.10).
Salient Features. The salient features of the organic cellular reserve materials present in the prokaryotes are as stated under :
(1) Poly-β-hydroxybutyric acid. It is found exclusively in the prokaryotes and invariably ca-ters as an equivalent of lipoidal content duly stored in the eukaryotic cells. It is observed in several species of Azotobacter, bacilli, and pseudomonads. Interestingly, certain specific or-ganisms viz., purple bacteria has the ability to synthesize even two types of reserve materi-als (e.g., glycogen and poly-β-hydroxybutyrate) simultaneously.
(a) Visibility — These organic cellular reserve materials are found to be deposited almost uniformly very much within the cytoplasm ; however, they may not be detected under a light microscope unless and until these are duly stained.
(b) Cellular content — The actively ‘growing cells’ do have these reserve materials present in rather small quantum in the cellular content ; whereas, they get usually accumulated exclusively in the C-rich culture medium under the influence of restricted amounts of nitrogen.
(c) Availability — These reserve materials may sometimes represent even upto 50% of the total cellular content on dry weight basis.
(d) Utility — These reserve materials are fully utilized when the prevailing cells are ad-equately provided with a suitable source of N and the growth is resumed subsequently.
(2) Glycogen and Starch — It has been duly established that the synthesis of glycogen and starch is usually accomplished via a proven mechanism for storing C in a form which is osmotically inert ; whereas, in the particular instance of poly-β-hydroxybutyric acid it pre-cisely designates a method of neutralizing an acidic metabolite.
(3) Cyanophycine (a copolymer of arginine and aspartic acid) :
In general, prokaryotes fail to store particularly the organic nitrogenous materials, but the blue-green bacteria is expected which essentially accumulate a nitrogenous reserve material termed as cyanophycine. It invariably represents as much as 8% of cellular dry weight; and may be regarded as a copolymer of arginine and aspartic acid.
(4) Volutin (metachromatic) Granules. A plethora of prokaryotes acquire more and more of volutin granules that may be stained meticulously with a ‘basic dye’, for instance : methyl-ene blue. In fact, these prokaryotes appear as red on being stained with a ‘blue-dye’. Impor-tantly, the prevailing metachromatic nature of the ensuing ‘red complex’ is on account of the very presence of a substantial quantum of ‘inorganic phosphates’. Evidently, the actual accumulation of these substances in the prokaryotes takes place under critical parameters of starvation specifically during ‘sulphate starvation’. It has been observed that these instantly generated volutin granules disappear as soon as the cells are adequately made available with a ‘sulphur source’, and subsequently the phosphate moiety [PO43–] is incorporated strategi-cally into the nucleic acids i.e., DNA and RNA. From the above statement of facts one may vividly infer that the ‘volutin granules’ definitely represent particularly the ‘intracellular phosphate reserve’ when the desired nucleic acid synthesis fails to materialize.
(5) Sulphur Bacteria [e.g., photosynthetic purple sulphur bacteria ; and filamentous non-photosynthetic bacteria (viz., Baggiatoa and Thiothrix)]. The aforementioned two sulphur bacteria specifically help in the accumulation of ‘Sulphur’ transiently in the course of hydro-gen sulphide [H2S] oxidation.
(6) Thylakoids. These are solely present in the blue-green bacteria and are intimately involved in the phenomenon of photosynthesis. Besides, there are three prominent structures, namely : gas vesicles, chlorobium vesicles, and carboxysomes, that are critically bound by non-unit membranes have been reported to be present in certain photosynthetic organisms.
(7) Ribs. There are several aquatic prokaryotes essentially containing gas vacuoles that are intimately engaged in counter-balancing the prevailing gravitational pull appreciably. On being examined under a ‘light microscope’ the ensuing gas vacuoles do look like dense refractile structure having a distinct irregular peripheral boundary. Importantly, with a cer-tain surge in the hydrostatic built-up pressure the existing gas vacuoles collapse thereby the cells lose their buoyancy eventually. Precisely, each gas vesicle more or less has an appear-ance very much akin to a ‘hollow cylinder’ having an approximate diameter of 75 nm with distinct conical ends, and a length ranging between 200 and 1000 nm. These conglomerates of gas vesicles are usually surrounded by a layer of protein approx. 2 mm thick. These structures do possess several bands consisting of regular rows of subunits that almost run perpendicular to the axis, and are termed as ‘ribs’. The ribs are found to be impermeable to water.
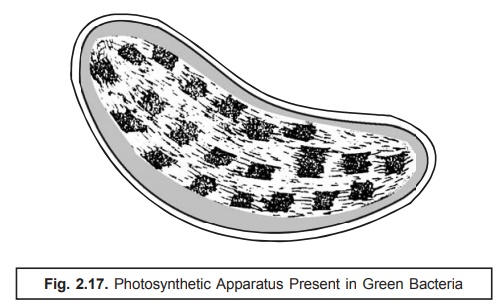
(8) Photosynthetic Apparatus. The photosynthetic apparatus present specifically in the pho-tosynthetic green bacteria (chlorobium) possesses a distinct strategic intracellular loca-tion. It is usually bound by a series of cigar-shaped vesicles arranged meticulously in a corticle-layer which immediately underlies the cell membrane as illustrated in Fig. 2.17. Interestingly, these structures have a width nearly 50 nm, length varying between 100–150 nm and are delicating enclosed within a single layered membrane of thickness ranging be-tween 3–5 nm. They essentially and invariably contain the ‘photosynthetic pigments’.
(9) Carboxysomes. It has been amply demonstrated that a good number of photosynthetic and chemolithotrophic organisms, namely : blue-green bacteria, purple bacteria, and thiobacilli essentially comprise of polyhedral structures having a width of 50–500 nm and carefully surrounded by a single layer of membrane having a thickness of 3.5 nm approximately. These characteristic structures are known as carboxysomes. They are found to consist of certain key enzymes that are closely associated with and intimately involved in the critical fixation of carbon dioxide [CO2], such as : carboxy dismutase ; and thus, represent the precise and most probable site of CO2 fixation in the photosynthetic as well as chemolithotrophic organisms.
Related Topics