Humoral Immune Effector Functions
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Immunology
The humoral immune response is mediated by the initial antibody–antigen interaction, but with the different antibody classes offering a range of effector functions. The effector functions of antibodies include those described below.
HUMORAL
IMMUNE EFFECTOR FUNCTIONS
The humoral immune response is mediated by
the initial antibody–antigen interaction, but with the different antibody
classes offering a range of effector functions. The effector functions of
antibodies include those described below.
1) Cognitive Function On B-Lymphocyte Cell Surface
Antibody on the surface of naive or memory
B-lymphocytes serves to recognize and bind specific antigen; IgM serves this
main cognitive function. It exists as a pentamer of five monomer units with an
antigen valency of 10 and is extremely efficient at binding antigen. IgD
appears to function mainly on the surface of B-lymphocytes and may also
contribute to cognition in some way.
2) Neutralization Of Antigen By Secreted Antibody
Secreted antibody, in particular IgG, IgA
and IgM, can bind antigen and sterically hinder the interaction of toxins,
viruses, bacteria, etc. with host cell surfaces. In the circulatory and
interstitial fluids IgG (which exists as a monomer with an antigen valency of
2) is the main antibody that fulfils this role in the secondary response, while
IgM is the main antibody produced in the primary response. IgA has specific
roles in mucosal immunity.
3) Opsonization Of Antigen
Secreted antibody, in particular IgG,
opsonizes antigenic material and in doing so promotes association (e.g. through
Fcg receptors) of the antigenic material with phagocyte membranes. Occupancy of
the Fc receptor by the antibody also serves to activate a phagocyte’s killing
mechanisms.
4) Mucosal Immunity
Mucosal immunity involves the interaction
of antibody with antigen at mucosal surfaces such as those of the
gastrointestinal tract, lung or urogenital tract. The major antibody of the
mucosal lining fluid is IgA, which exists as a dimer of two monomer units
(antigen valency of 4). IgA is actively secreted across mucosal epithelium into
the lining fluid; it will neutralize antigen and may also serve as an opsonin.
IgA is also present in secretions such as tears, saliva, etc. but it has a
limited role in systemic immunity.
5) Antibody-Dependent Cell Cytotoxicity (ADCC)
Through specific binding to antigen on the
surface of membranes perceived as ‘foreign’, e.g. microbial cells or host cells
virally infected or otherwise transformed, antibody can direct (through its Fc
domains) the close association of ‘killing’ cells, such as neutrophils,
eosinophils, NK cells and even cytotoxic T-lymphocytes, with the ‘foreign’
membrane. This close association depends on the antibody’s Fc domain binding to
the respective Fc receptor present on the surface membrane of the ‘killing’
cell. Such close proximity to the ‘foreign’ cell enables the efficient and
targeted release of cytotoxic molecules into the extracellular environment. IgG
is the main antibody of systemic body fluids and is an important mediator of
ADCC, although IgE and IgA may undertake this role in certain circumstances,
e.g. against certain parasites IgE directs ADCC mediated by eosinophils.
6) Immediate Hypersensitivity
Mast cells express high-affinity receptors
(Fce) that bind the Fc domain of IgE antibodies. In the absence of antigen
these receptors are occupied by the IgE monomer (antigen valency of 2) secreted
previously from plasma cells. In this circumstance the IgE molecules are
serving a cognitive function which, on appropriate antigen binding, results in
aggregation of the membrane-bound IgE and causes immediate mast cell
degranulation and release of inflammatory mediators. Mast cells possess in
their membranes IgE monomers able to recognize different antigenic epitopes.
This contrasts with each single B-lymphocyte, which possesses IgM antibody on
its surface membrane that performs a cognitive function but is capable of
recognizing only a single epitope specificity.
7) Neonatal Immunity
The neonate lacks the ability to mount a
full immunological response; accordingly, maternal IgG is transported across
the placenta late in pregnancy and is also absorbed across the gastrointestinal
tract from breast milk. Maternal IgA secreted into breast milk will also
provide mucosal protection for the neonate.
8) Activation Of The Classical Complement Pathway
A complement cascade similar to that of
the alternative pathway can be activated through specific antibody–antigen
interactions. The antibodies that activate the classical complement pathway are
IgM and IgG.
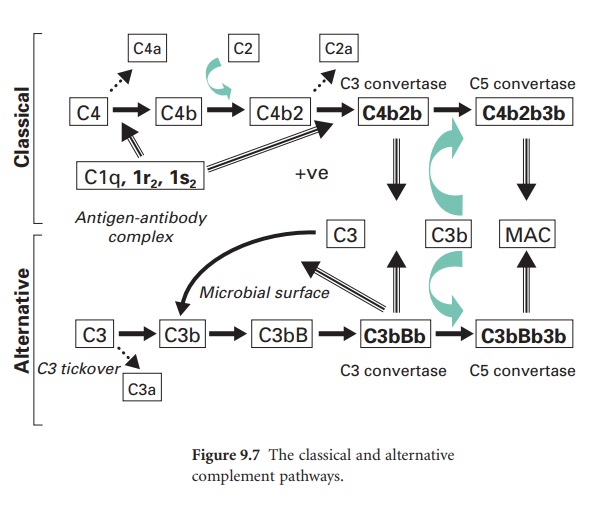
Key steps in the activation of the
classical pathway are shown in Figure 9.7, where this pathway is also compared
to the alternative pathway. In the classical pathway the initiating step is the
specific binding of IgG or IgM to antigen. Once this occurs, a complement
protein termed C1 (which comprises a single C1q subunit, two C1r subunits and
two C1s subunits) binds to adjacent Fc domains in the antibody–antigen complex.
This binding of C1 activates the catalytic activity of the C1r subunits, and in
turn the C1s subunits. The activated C1s subunits cleave C4 into C4b and C4a;
the latter can diffuse away and serve as a leucocyte activator. The C4b
covalently associates with the antibody–antigen complex on the surface of a
microbial membrane and can serve as an opsonin. A further complement protein,
C2, binds to this membrane complex to give C4b2. The C1s subunit then
enzymically cleaves the bound C2a to generate on the membrane a new complex termed
C4b2b, which is the C3 convertase of the classical pathway. (In some texts the
C2a is referred to as the larger subunit remaining with the membrane while C2b
is the smaller subunit that diffuses away.)
This C3 convertase molecule is distinct
from that within the alternative pathway, but it is from this point onwards
that parallels can be drawn between the two cascades.
The host proteins that serve key regulatory functions within the alternative pathway (DAF, CR1 factor I, CD59) also serve similar functions within the classical pathway. However, in contrast to the alternative pathway the activation step in the classical pathway requires specific antibody–antigen interactions. In this context the C1 protein can only become catalytically active when it is bound to at least two adjacent Fc domains. In the case of the IgG and IgM molecules the Fc domains will only align adjacent to each other when the corresponding Fab domains bind antigen. Further, when C1 is free in the circulation it is bound to a protein termed C1 inhibitor (C1-INH) which prevents any possible activation of C1 in the absence of antibody. Once C1 binds to adjacent Fc domains within an antibody–antigen complex C1-INH is displaced.
The functions of the classical complement
pathway are similar to those described for the alternative pathway, i.e.
opsonization, leucocyte activation and membrane lysis of target cells. The
classical pathway can additionally lead to complement protein deposition on
insoluble antibody– antigen immune complexes circulating within blood, and in
doing so promote the clearance of such potentially harmful complexes by Kupffer
cells of the liver. The presence of two complement pathways provides for rapid
(alternative) and specific (classical) activation of a key defence mechanism,
and offers greater protection against the development of microbial resistance
mechanisms.
Related Topics
