Manifestation of Disease
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Principles Of Microbial Pathogenicity And Epidemiology
Once established, the course of a bacterial infection can proceed in a number of ways. These can be related to the relative ability of the organism to penetrate and invade surrounding tissues and organs.
MANIFESTATION OF DISEASE
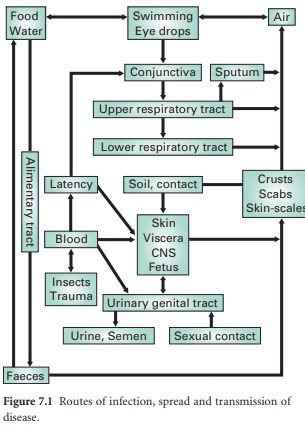
Once established, the
course of a bacterial infection can proceed in a number of ways. These can be
related to the relative ability of the organism to penetrate and invade
surrounding tissues and organs. The vast majority of pathogens, being unable to
combat the defences of the deeper tissues, consolidate further on the
epithelial surface. Others, including most viruses, penetrate the epithelial
layers, but no further, and can be regarded as partially invasive. A small
group of pathogens are fully invasive. These permeate the subepithelial tissues
and are circulated around the body to initiate secondary sites of infection
remote from the initial portal of entry (Figure 7.1).
Other groups of
organisms may cause disease through ingestion by the victim of substances
produced during microbial growth on foods. Such diseases may be regarded as
intoxications rather than as infections and are considered later. Treatment in
these cases is usually an alleviation of the harmful effects of the toxin
rather than elimination of the pathogen from the body.
1) NONINVASIVE PATHOGENS
Bordetella pertussis (the aetiological agent of whooping cough) is probably the best described
of these pathogens. This organism is inhaled and rapidly localizes on the
mucociliary blanket of the lower respiratory tract. This localization is very
selective and is thought to involve agglutinins on the organism’s surface.
Toxins produced by the organism inhibit ciliary movement of the epithelial
surface and thereby prevent removal of the bacterial cells to the gut. A high
molecular weight exotoxin is also produced during the growth of the organism
which, being of limited diffusibility, pervades the subepithelial tissues to
produce inflammation and necrosis. Corynebacterium diphtheriae (the causal organism of diphtheria) behaves similarly, attaching itself to the
epithelial cells of the respiratory tract. This organism produces a diffusible
toxin of low molecular weight, which enters the blood circulation and brings
about a generalized toxaemia.
In the gut, many
pathogens adhere to the gut wall and produce their effect via toxins that
pervade the surrounding gut wall or enter the systemic circulation. V. cholerae and some enteropathic E. coli strains localize on the gut wall
and produce toxins that increase vascular permeability. The end result is a
hypersecretion of isotonic fluids into the gut lumen, acute diarrhoea and, as a
consequence, dehydration that may be fatal in young or elderly people. In all
these instances, binding to epithelial cells is not essential but increases permeation
of the toxin and prolongs the presence of the pathogen.
2) PARTIALLY INVASIVE PATHOGENS
Some bacteria, and most
viruses, are able to attach to the mucosal epithelia and then penetrate rapidly
into the epithelial cells. These organisms multiply within the protective environment
of the host cell, eventually killing it and inducing disease through erosion
and ulceration of the mucosal epithelium. Typically, members of the genera Shigella and Salmonella utilize such mechanisms in infections of the
gastrointestinal tract. These bacteria attach to the epithelial cells of the
large and small intestines, respectively, and, following their entry into these
cells by induced pinocytosis, multiply rapidly and penetrate laterally into
adjacent epithelial cells. The mechanisms for such attachment and movement are
unknown but involve a transition from a nonmotile to motile phenotype. Some species
of salmonellae produce, in addition, exotoxins that induce diarrhoea (section
6.1.4). There are innumerable serotypes of Salmonella,
which are primarily parasites of animals but are important to humans in that
they colonize farm animals such as pigs and poultry and ultimately infect foods
derived from them. Salmonella food
poisoning (salmonellosis), therefore, is commonly associated with inadequately
cooked meats, eggs and also with cold meat products that have been incorrectly
stored following contact with the uncooked product. Dependent upon the severity
of the lesions induced in the gut wall by enteric pathogens, red blood cells
and phagocytes pass into the gut lumen, along with plasma, and cause the
classic ‘bloody flux’ of bacillary dysentery. Similar erosive lesions are
produced by some enteropathic strains of E.
coli.
Viral infections such as influenza and the ‘common cold’ (in reality 300–400 different strains of rhinovirus) infect epithelial cells of the respiratory tract and nasopharynx, respectively. Release of the virus, after lysis of the host cells, is to the void rather than to subepithelial tissues. The residual uninfected epithelial cells are rapidly infected, resulting in general degeneration of the tracts. Such damage not only predisposes the respiratory tract to infection with opportunist pathogens such as N. meningitidis and Haemophilus influenzae but also causes the associated fever.
3) FULLY INVASIVE PATHOGENS
Invasive pathogens
either aggressively invade the tissues surrounding the primary site of
infection (active spread) or are passively transported around the body in the
blood, lymph, cerebrospinal, axonal or pleural fluids (passive spread). Some,
especially aggressive organisms, move both passively and actively, setting up
multiple, expansive secondary sites of infection in various organs.
a) Active spread
Active spread of
microorganisms through normal subepithelial tissues is difficult in that the
gel like nature of the intercellular materials physically inhibits bacterial movement.
Induced death and lysis of the tissue cells produces, in addition, a highly viscous
fluid, partly due to undenatured DNA. Physical damage, such as wounds, is
rapidly sealed with fibrin clots, thereby reducing the effective routes for
spread of opportunist pathogens. Organisms such as Strep. pyogenes, Cl. perfringens and, to some extent, the staphylococci,
are able to establish themselves in tissues by virtue of their ability to
produce a wide range of extracellular enzyme toxins. These are associated with
killing of tissue cells, degradation of intracellular materials and mobilization
of nutrients. A selection of such toxins will be considered briefly.
Haemolysins are produced by most of the pathogenic staphylococci and streptococci. They have a lytic effect on red
blood cells, releasing iron containing nutrients.
Fibrinolysins are produced by both
staphylococci (staphylokinase) and
streptococci (streptokinase). These toxins indirectly activate plasminogen and
so dissolve fibrin clots that the host forms around wounds and lesions to seal
them. The production of fibrinolysins therefore increases the likelihood of the
infection spreading. Streptokinase may be employed clinically in conjunction
with streptodornase in the treatment of thrombosis.
Collagenases and hyaluronidases are produced by most of the aggressive invaders of tissues. These are able to dissolve
collagen fibres and the hyaluronic acids that function as intercellular
cements; this causes the tissues to break up and produce oedematous lesions.
Phospholipases are produced by
organisms such as Cl. perfringens (αtoxin). These toxins kill tissue cells by hydrolysing the phospholipids that are
present in cell membranes.
Amylases, peptidases and deoxyribonucleases mobilize many
nutrients that are released from lysed cells. They also decrease the viscosity
of fluids present at the lesion by depolymerization of their biopolymer
substrates.
Organisms possessing the
above toxins, particularly those also possessing leucocidins, are likely to cause expanding oedematous lesions at
the primary site of infection. In the case of Cl. perfringens, a soil microorganism that has become adapted to a
saprophytic mode of life, infection arises from an accidental contamination of
deep wounds when a process similar to that seen during the decomposition of a
carcass ensues (gangrene). This organism is most likely to spread through
tissues when blood circulation, and therefore oxygen tension, in the affected
areas is minimal.
Abscesses formed by
streptococci and staphylococci can be deep seated in soft tissues or associated with infected
wounds or skin lesions; they become localized through the deposition of fibrin
capsules around the infection site. Fibrin deposition is partly a response of
the host tissues, but is also partly a function of enzyme toxins such as coagulase . Phagocytic white blood cells
can migrate into these abscesses in large numbers to produce significant
quantities of pus. Such pus, often carrying the infective pathogen, might be
digested by other phagocytes in the late stages of the infection or discharged
to the exterior or to the capillary and lymphatic network. In the latter case,
blocked capillaries might serve as sites for secondary lesions. Toxins
liberated from the microorganisms during their growth in such abscesses can
freely diffuse to the rest of the body to set up a generalized toxaemia.
S. enterica serovar Typhi, S. enterica serovar
Paratyphi and S. enterica serovar Typhimurium are serotypes of Salmonella (section 4.2) that are not only
able to penetrate into intestinal epithelial cells and produce exotoxins, but are
also able to penetrate beyond into subepithelial tissues. These organisms
therefore produce a characteristic systemic disease (typhoid and enteric
fever), in addition to the usual symptoms of salmonellosis. Following recovery
from such infection the organism is commonly found associated with the
gallbladder. In this state, the recovered person will excrete the organism and
become a reservoir for the infection of others.
b) Passive spread
When invading
microorganisms have crossed the epithelial barriers they will almost certainly
be taken up with lymph in the lymphatic ducts and be delivered to filtration
and immune systems at the local lymph nodes. Sometimes this serves to spread
infections further around the body. Eventually, spread may occur from local to
regional lymph nodes and thence to the bloodstream. Direct entry to the
bloodstream from the primary portal of entry is rare and will only occur when
the organism damages the blood vessels or if it is injected directly into them.
This might be the case following an insect bite or surgery. Bacteraemia such as
this will often lead to secondary infections remote from the original portal of
entry.
Related Topics
