Plant Hormones and Growth Regulators
| Home | | Pharmacognosy |Chapter: Pharmacognosy and Phytochemistry : Cultivation, Collection and Processing of Herbal Drugs
Plant hormones (phytohormones) are physiological inter-cellular messengers that control the complete plant lifecycle, including germination, rooting, growth, flowering, fruit ripening, foliage and death. In addition, plant hormones are secreted in response to environmental factors such as excess of nutrients, drought conditions, light, temperature and chemical or physical stress.
PLANT HORMONES AND GROWTH REGULATORS
Plant hormones (phytohormones) are physiological
inter-cellular messengers that control the complete plant lifecycle, including
germination, rooting, growth, flowering, fruit ripening, foliage and death. In
addition, plant hormones are secreted in response to environmental factors such
as excess of nutrients, drought conditions, light, temperature and chemical or
physical stress. So, levels of hormones will change over the lifespan of a
plant and are dependent upon season and environment.
The term ‘plant growth factor’ is usually employed for plant
hormones or substances of similar effect that are administered to plants.
Growth factors are widely used in industrialized agriculture to improve
productivity. The application of growth factors allows synchronization of plant
development to occur. For instance, ripening fruits can be controlled by
setting desired atmospheric ethylene levels. Using this method, fruits that are
separated from their parent plant will still respond to growth factors;
allowing commercial plants to be ripened in storage during and after
transportation. This way the process of harvesting can be run much more
efficiently and effectively. Other applica-tions include rooting of seedlings
or the suppression of rooting with the simultaneous promotion of cell division
as required by plant cell cultures. Just like with animal hormones, plant
growth factors come in a wide variety, producing different and often
antagonistic effects. In short, the right combination of hormones is vital to
achieve the desired behavioural characteristics of cells and the produc-tive
development of plants as a whole. The plant growth regulators are classified
into synthetic and native. The synthetic regulators are also known as exogenous
regulators and the native are called the endogenous,
Five major classes of plant hormones are mentioned: auxins,
cytokinins, gibbereilins, abscisic acid and ethylene. However as research
progresses, more active molecules are being found and new families of
regulators are emerging; one example being polyamines (putrescine or
spermidine). Plant growth regulators have made the way for plant tissue culture
techniques, which were a real boon for mankind in obtaining therapeutically
valuable secondary metabolites.
Auxins
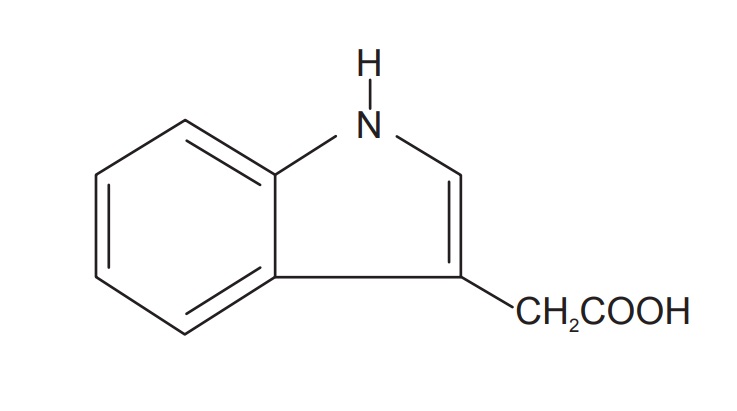
The term auxin is derived from the Greek word auxein which
means to grow. Generally compounds are considered as auxins if they are able to
induce cell elongation in stems and otherwise resemble indoleacetic acid (the
first auxin isolated) in physiological activity. Auxins usually affect other
processes in addition to cell elongation of stem cells but this characteristic
is considered critical of all auxins and thus ‘helps’ define the hormone.
Auxins were the first plant hormones discovered. Charles
Darwin was among the first scientists to pool in plant hormone research. He
described the effects of light on movement of canary grass coleoptiles in his
book ‘The Power of Movement in Plants’ presented in 1880. The coleoptile is a
specialized leaf originating from the first node which sheaths the epicotyl in
the plants seedling stage protecting it until it emerges from the ground. When
unidirectional light shines on the coleoptile, it bends in the direction of the
light. If the tip of the coleoptile was covered with aluminium foil, bending
would not occur towards the unidirectional light. However if the tip of the
coleoptile was left uncovered but the portion just below the tip was covered,
exposure to unidirectional light resulted in curvature toward the light.
Darwin’s experiment suggested that the tip of the coleoptile was the tissue
responsible for perceiving the light and producing some signal which was
transported to the lower part of the coleoptile where the physiological
response of bending occurred. When he cut off the tip of the coleoptile and
exposed the rest of the coleoptile to unidirectional light curvature did not
occur confirming the results of his experiment.
Salkowski (1885) discovered indole-3-acetic acid (IAA) in
fermentation media. The isolation of the same product from plant tissues would
not be found in plant tissues for almost 50 years. IAA is the major auxin
involved in many of the physiological processes in plants. Fitting in 1907 put
his efforts in studying signal transaction by making incisions on the light or
dark side of the plant. He failed because the signal was capable of crossing or
going around the incision, In 1913, modification was made in Fitting’s
experiment by Boysen-Jensen, in that they inserted pieces of mica to block the
transport of the signal and showed that transport of auxin toward the base
occurs on the dark side of the plant as opposed to the side exposed to the
unidirectional light. In 1918, Paal confirmed Boysen-Jensen’s results by
cutting off coleoptile tips in the dark, exposing only the tips to the light,
replacing the coleoptile tips on the plant but off centered to one side or the
other. Results showed that whichever side was exposed to the coleoptile,
curvature occurred toward the other side. Soding 1925, followed Paal’s idea and
showed that if tips were cut off there was a reduction in growth but if they
were cut off and then replaced growth continued to occur.
In 1926, Fritz Went reported a plant growth substance,
isolated by placing agar blocks under coleoptile tips for a period of time then
removing them and placing them on decapitated Avena stems. After placement over
the agar, the stems resumed growth. In 1928, again Went developed a method of
quantifying this plant growth substance. His results suggested that the
curvatures of stems were proportional to the amount of growth substance in the
agar. This test was called the avena curvature test. Much of our current
knowledge of auxin was obtained from its applications. It was Went’s work,
which had a great influence in stimulating plant growth substance research. He
is often credited with dubbing the term auxin but it was actually Kogl and
Haagen-Smit who purified the compound auxentriolic acid (auxin A) from human
urine in 1931. Later Kogl isolated other compounds from urine which were
similar in structure and function to auxin A. One of which was indole-3 acetic
acid (IAA) initially discovered by Salkowski in 1885. In 1954 a committee of
plant physiologists was set up to characterize the group auxins.
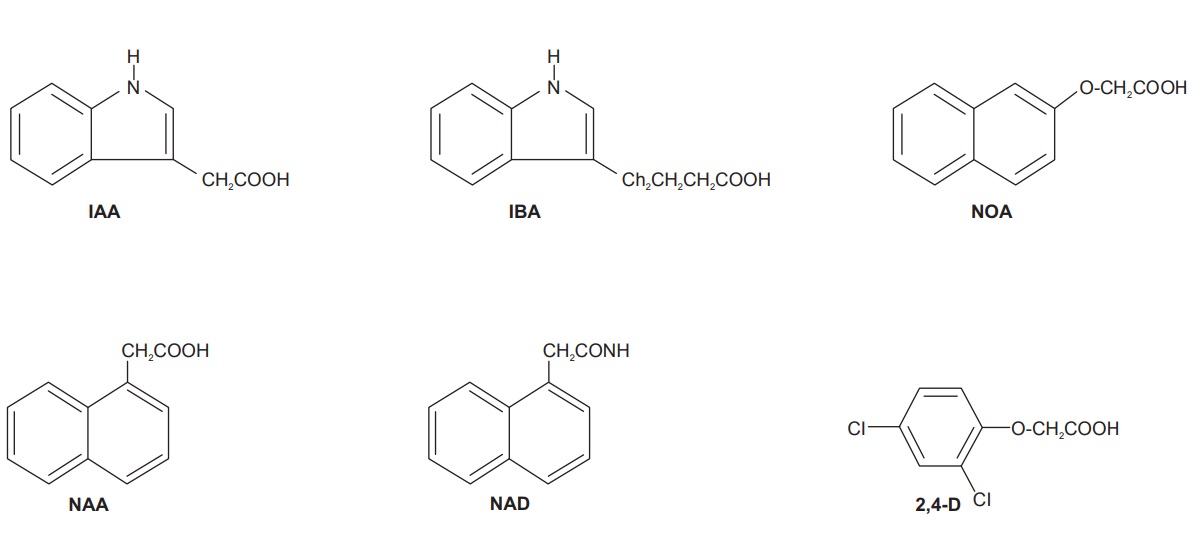
Indole acetic acid (IAA) is the principle natural auxin and
other natural auxins are indole-3-acetonitrile (IAN), phenyl acetic acid and
4-chloroindole-3-acetic acid. The exogenous or synthetic auxins are
indole-3-butyric acid (IBA), α-napthyl acetic acid (NAA), 2-napthyloxyacetic
acid (NOA), 1-napthyl acetamide (NAD), 5-carboxymeth-yl-N, N-dimethyl
dithiocarbamate, 2,4-dichlorophenoxy acetic acid (2,4-D), etc.
Production
and occurrence
Produced in shoot and root meristematic tissue, in young
leaves, mature root cells and small amounts in mature leaves. Transported
throughout the plant parts and the production of IAA will be more in day time.
It is released by all cells when they are experiencing conditions which would
normally cause a shoot meristematic cell to produce auxin. Ethylene has direct
or indirect action over to enhance the synthesis auxin.
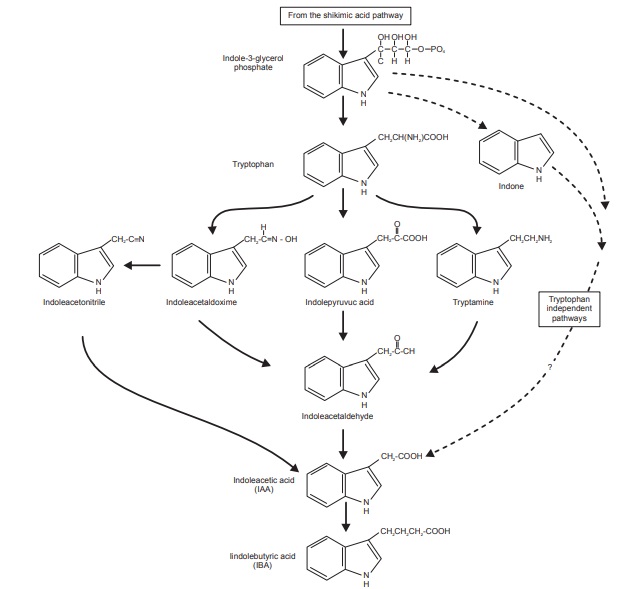
IAA is chemically similar to the amino acid tryptophan which
is generally accepted to be the molecule from which IAA is derived. Three
mechanisms have been suggested to explain this conversion:
· Tryptophan is converted to
indolepyruvic acid through a transamination reaction. Indolepyruvic acid is
then converted to indoleacetaldehyde by a decarboxylation reaction. The final
step involves oxidation of indoleac-etaldehyde resulting in indoteacetic acid.
· Tryptophan undergoes decarboxylation
resulting in tryptamine. Tryptamine is then oxidized and deaminated to produce
indoleacetaldehyde. This molecule is further oxidized to produce indoleacetic
acid.
· IAA can be produced via a
tryptophan-independent mechanism. This mechanism is poorly understood, but has
been proven using tip (-) mutants. Other experiments have shown that, in some
plants, this mechanism is actually the preferred mechanism of IAA biosynthesis.
The enzymes responsible for the biosynthesis of IAA are most
active in young tissues such as shoot apical meristems and growing leaves and
fruits. These are the same tissues where the highest concentrations of IAA are
found. One way plants can control the amount of IAA present in tissues at a
particular time is by controlling the biosynthesis of the hormone. Another
control mechanism involves the production of conjugates which are, in simple
terms, mol-ecules which resemble the hormone but are inactive. The formation of
conjugates may be a mechanism of storing and transporting the active hormone.
Conjugates can be formed from IAA via hydrolase enzymes. Conjugates can be
rapidly activated by environmental stimuli signaling a quick hormonal response.
Degradation of auxin is the final method of controlling auxin levels. This
process also has two proposed mechanisms outlined below:
The oxidation of IAA by oxygen resulting in the loss of the
carboxyl group and 3-methyleneoxindole as the major breakdown product. IAA
oxidase is the enzyme which catal-yses this activity. Conjugates of IAA and
synthetic auxins such as 2,4-D can not be destroyed by this activity.
C-2 of the heterocyclic ring may be oxidized resulting in
oxindole-3-acetic acid. C-3 may be oxidized in addition to C-2 resulting in
dioxindole-3-acetic acid. The mechanisms by which biosynthesis and degradation
of auxin molecules occur are important to future agricultural applications.
Information regarding auxin metabolism will most likely lead to genetic and
chemical manipulation of endogenous hormone levels resulting in desirable
growth and differ-entiation of important plant species.
Functions
of auxin
· Stimulates cell elongation.
· The auxin supply from the apical bud
suppresses growth of lateral buds. Apical dominance is the inhibiting
influ-ence of the shoot apex on the growth of axillary buds. Removal of the
apical bud results in growth of the axillary buds. Replacing the apical bud
with a lanolin paste containing IAA restores the apical dominance. The
mechanism involves another hormone - ethylene. Auxin (IAA) causes lateral buds
to make ethylene, which inhibits growth of the lateral buds.
· Differentiation of vascular tissue
(xylem and phloem) is stimulated by IAA.
· Auxin stimulates root initiation on
stem cuttings and lateral root development in tissue culture (adventitious
rooting).
· Auxin mediates the tropistic
response of bending in response to gravity and light (this is how auxin was
first discovered).
· Auxin has various effects on leaf
and fruit abscission, fruit set, development, and ripening, and flowering,
depending on the circumstances.
Cytokinins
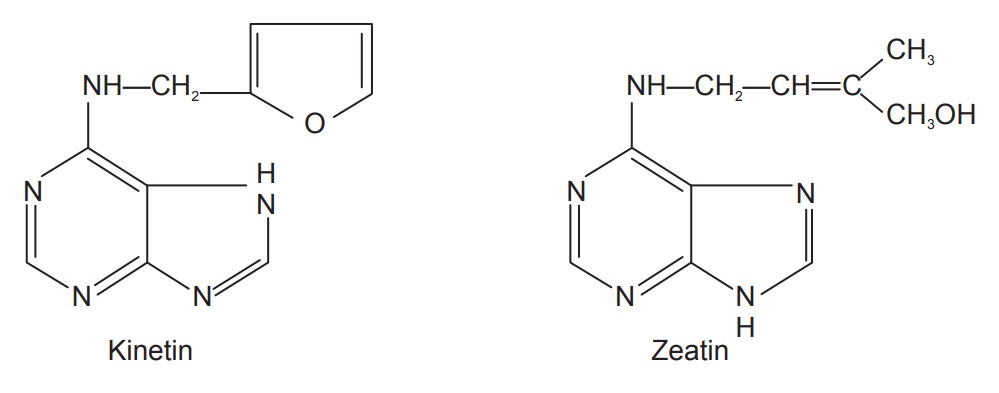
Cytokinins are compounds with a structure resembling adenine
which promote cell division and have other similar functions to kinetin. They
also regulate the pattern and frequency of organ production as well as position
and shape. They have an inhibitory effect on senescence. Kinetin was the first
cytokinin identified and so named because of the compounds ability to promote
cytokinesis (cell division). Though it is a natural compound, it is not made in
plants, and is therefore usually considered a ‘synthetic’ cytokinin. The common
naturally occurring cytokinin in plants today is called zeatin which was
isolated from corn.
Cytokinin have been found in almost all higher plants as
well as mosses, fungi, bacteria, and also in many prokary-otes and eukaryotes.
There are more than 200 natural and synthetic cytokinins identified. Cytokinin
concentrations are more in meristematic regions and areas of continuous growth
potential such as roots, young leaves, developing fruits, and seeds.
Haberlandt (1913) and Jablonski and Skoog (1954) identified
that a compound found in vascular tissues had the ability to stimulate cell
division. In 1941, Johannes van Overbeek discovered that the milky endosperm
from coconut and other various species of plants also had this ability. The
first cytokinin was isolated from herring sperm in 1955 by Miller and his
associates. This compound was named kinetin because of its ability to promote
cytokine-sis (cell division). The first naturally occurring cytokinin was
isolated from corn in 1961 by Miller and it was later called zeatin. Since that
time, many more naturally occur-ring cytokinins have been isolated and the
compound was common to all plant species in one form or another.
The naturally occurring cytokinins are zeatin, N6
dim-ethyl amino purine, isopentanyl aminopurine. The syn-thetic cytokinins are
kineatin, adenine, 6-benzyl adenine benzimidazole and N, N’-diphenyl urea.
Production
and occurrence
Produced in root and shoot meristematic tissue, in mature
shoot cells and in mature roots in small amounts. If is rapidly transported in
xylem stream. Peak production occurs in day time and their activity is reduced
in plants suffering drought. It is directly or indirectly induced by high
levels of Gibberlic acid.
Cytokinin is generally found in meristematic regions and
growing tissues. They are believed to be synthesized in the roots and
translocated via the xylem to shoots. Cytokinin biosynthesis happens through
the biochemical modification of adenine. They are synthesized by following
pathway.
A product of the mevalonate pathway called isopentyl
pyrophosphate is isomerized. This isomer can then react with adenosine
monophosphate with the aid of an enzyme called isopentenyl AMP synthase. The
result is isopentenyl adenosine-5’-phosphate (isopentenyl AMP). This product
can then be converted to isopentenyl adenosine by removal of the phosphate by a
phosphatase and further converted to isopentenyl adenine by removal of the
ribose group. Isopentenyl adenine can be converted to the three major forms of
naturally occurring cytokinins.
Other pathways or slight alterations of this one probably
lead to the other forms. Degradation of cytokinins occurs largely due to the
enzyme cytokinin oxidase. This enzyme removes the side chain and releases
adenine. Derivatives can also be made but the difficulties are with pathways,
which are more complex and poorly understood.
Functions
of cytokinin
· Stimulate cell division
(cytokinesis).
· Stimulate morphogenesis (shoot
initiation/bud forma-tion) in tissue culture.
· Stimulate the growth of lateral (or
adventitious) buds-release of apical dominance.
· Stimulate leaf expansion resulting
from cell enlarge-ment.
· May enhance stomatal opening in some
species (Figure 6.2).
· Promotes the conversion of
etioplasts into chloroplasts via stimulation of chlorophyll synthesis.
· Stimulate the dark-germination of
light-dependent seeds.
· Delays senescence.
· Promotes some stages of root
development.

Effect of cytokinin on stomatal opening
Ethylene
Ethylene has been used in practice since the ancient times,
where people would use gas figs in order to stimulate ripen-ing, burn incense
in closed rooms to enhance the ripening of pears. It was in 1864, that leaks of
gas from street lights showed stunting of growth, twisting of plants, and
abnormal thickening of stems. In 1901, a Russian scientist named Dimitry
Neljubow showed that the active component was ethylene. Doubt 1917, discovered
that ethylene stimulated abscission. In 1932 it was demonstrated that the
ethylene evolved from stored apple inhibited the growth of potato shoots
enclosed with them. In 1934 Gane reported that plants synthesize ethylene. In
1935, Crocker proposed that ethylene was the plant hormone responsible for
fruit ripening as well as inhibition of vegetative tissues. Ethylene is now
known to have many other functions as well.
Production
and occurrence
Production is directly induced by high levels of Auxin, root
flooding and drought. It is found in germinating seeds and produced in nodes of
stems, tissues of ripening fruits, response to shoot environmental, pest, or
disease stress and in senescent leaves and flowers. Light minimizes the
production of ethylene. It is released by all cells when they are experiencing
conditions which would normally cause a mature shoot cell to produce ethylene.
Ethylene is produced in all higher plants and is produced
from methionine in essentially all tissues. Production of ethylene varies with
the type of tissue, the plant species, and also the stage of development. The
mechanism by which ethylene is produced from methionine is a three step
process. ATP is an essential component in the synthesis of ethylene from
methionine. ATP and water are added to methionine resulting in loss of the
three phosphates and S-adenosyl methionine (SAM). 1-amino-cyclopropane-l-carboxylic
acid synthase (ACC-synthase) facilitates the production of ACC from SAM. Oxygen
is then needed in order to oxidize ACC and produce ethylene. This reaction is
catalysed by an oxidative enzyme called ethylene forming enzyme. The control of
ethylene production has received considerable study. Study of ethylene has
focused around the synthesis promoting effects of auxin, wounding, and drought
as well as aspects of fruit-ripening. ACC synthase is the rate limiting step
for ethylene production and it is this enzyme that is manipulated in
biotechnology to delay fruit ripening in the ‘flavor saver’ tomatoes.
Functions
of ethylene
· Production stimulated during
ripening, flooding, stress, senescence, mechanical damage, infection.
· Regulator of cell death programs in
plants (apoptosis). Stimulates the release of dormancy.
· Stimulates shoot and root growth and
differentiation (triple response).
· Regulates ripening of climacteric
fruits.
· May have a role in adventitious root
formation. Stimulates leaf and fruit abscission.
· Flowering in most plants is
inhibited by ethylene. Mangos, pineapples and some ornamentals are stimu-lated
by ethylene.
· Induction of femaleness in dioecious
flowers. Stimulates flower opening.
· Stimulates flower and leaf
senescence.
Gibberellins
Unlike the classification of auxins which are classified on
the basis of function, gibberellins are classified on the basis of structure as
well as function. All gibberellins are derived from the ent-gibberellane
skeleton. The gibberellins are named GA1. GAn in order of
discovery. Gibberellic acid was the first gibberellin to be structurally
characterized as GA3. There are currently 136 GAs identified from
plants, fungi and bacteria.
They are a group of diterpenoid acids that functions as
plant growth regulators influencing a range of developmental processes in
higher plants including stem elongation, germination, dormancy, flowering, sex
expression, enzyme induction and leaf and fruit senescence. The origin of
research into gibberellins can be traced to Japanese plant pathologists who
were investigating the causes of the ‘bakanae’ (foolish seedling) disease which
seriously lowered the yield of rice crops in Japan, Taiwan and throughout the
Asian countries. Symptoms of the disease are pale yellow, elongated seedlings
with slender leaves and stunted roots. Severely diseased plants die whereas
plants with slight symptoms survive but produce poorly developed grain, or none
at all.
Bakanae is now easily prevented by treatment of seeds with
fungicides prior to sowing. In 1898 Shotaro Hori demonstrated that the symptoms
were induced by infection with a fungus belonging to the genus Fusarium,
probably Fusarium heterosporium Necs.
In 1912, Sawada suggested that the elongation in
rice-seedlings infected with bakanae fungus might be due to a stimulus derived
from fungal hyphae.
Subsequently, Eiichi Kurosawa (1926) found that culture
filtrates from dried rice seedlings caused marked elongation in rice and other
sub-tropical grasses. He concluded that bakanae fungus secretes a chemical that
stimulates shoot elongation, inhibits chlorophyll formation and suppresses root
growth.
Although there has been controversy among plant pathologists
over the nomenclature of bakanae fungus, in the 1930s, the imperfect stage of
the fungus was named Fusarium moniliforme
(Sheldon) and the perfect stage, was named
as Gibberella fujikuroi (Saw.) Wr. by
H.W. Wol-lenweber. The terms ‘Fujikuroi’ and ‘Saw’ in Gibberella fujikuroi (Saw.)
Wr. were derived from the names of two distinguished
Japanese plant pathologists, Yosaburo Fujikuro and Kenkichi Sawada.
In 1934, Yabuta isolated a crystalline compound from the
fungal culture filtrate that inhibited growth of rice seedlings at all
concentrations tested. The structure of the inhibitor was found to be
5-n-butylpicolinic acid or fusaric acid. The formation of fusaric acid in
culture filtrates was suppressed by changing the composition of the culture
medium. As a result, a noncrystalline solid was obtained from the culture
filtrate that stimulated the growth of rice seedlings. This compound was named
gibberellin by Yabuta.
In 1938, Yabuta and his associate Yusuke Sumiki finally
succeeded in crystallizing a pale yellow solid to yield gib-berellin A and
gibberellin B (The names were subsequently interchanged in 1941 and the
original gibberellin A was found to be inactive.) Determination of the
structure of the active gibberellin was hampered by a shortage of pure crystalline
sample. In the United States, the first research on gibberellins began after
the Second World War. In 1950, John E. Mitchell reported optimal fermentation
procedures for the fungus, as well as the effects of fungal extracts on the
growth of bean (Vicia faba)
seedlings. In Northern USDA Regional Research Laboratories in Peoria, large
scale fermentations were carried out with the purpose of producing pure
gibberellin A for agricultural uses but initial fermentations were found to be
inactive. Further researches were carried out by Sumiki in 1951, Stodola et
al., 1955, Curtis and Cross, 1954 regarding gibberellins and finally the
gibberllic acid was determined by its chemical and physical properties.
In 1955, members of Sumuki group, succeeded in sepa-rating
the methyl ester of gibberellin A into three compo-nents, from which
corresponding free acids were obtained and named gibberellins Al, A2, and A3.
Gibberellin A3 was found to be identical to gibberellic acid. In 1957,
Takahashi et al. isolated a new gibberellin named gibberellin A4 as a minor
component from the culture filtrate.
In the mid 1950s, evidence that gibberellins were naturally
occurring substances in higher plants began to appear in the literature.
Margaret Radley in the UK demonstrated the presence of gibberellin-like
substances in higher plants. In the United States, Bernard Phinney et al were
the first to report gibberellin-like substance in maize. This was followed by
the isolation of crystalline gibberellin Al, A5, A6 and A8 from runner bean (Phaseotus multiflorus). After 10 years
the number of gibberellins reported in the literature isolated from fungal and
plant origins rapidly increased. In 1968, J. MacMillan and N. Takahashi
concluded that all gibberellins should be assigned numbers as gibberellin A1-x,
irrespec-tive of their origin. Over the past 20 years using modern analytical
techniques many more gibberellins have been identified. At the present time the
number of gibberellins identified is 126.
Production
and occurrence
Produced in the roots, embryo and germinating seeds. The
level of gibberellins goes up in the dark when sugar cannot be manufactured and
will be reduced in the light. It is released in mature cells (particularly
root) when they do not have enough sugar and oxygen to support both themselves
and released by all cells when they are experiencing conditions which would
normally cause a mature root cell to produce GA.
Gibberellins are diterpenes synthesized from acetyl CoA via
the mevalonic acid pathway. They all have either 19 or 20 carbon units grouped
into either four or five ring systems. The fifth ring is a lactone ring as
shown in the structures above attached to ring A. Gibberellins are believed to
be synthesized in young tissues of the shoot and also the developing seed. It
is not clear whether young root tissues also produce gibberellins. There is
also some evidence that leaves may also contain them. The gibberellins are
formed through the pathway, three acetyl CoA molecules are oxi-dized by two
NADPH molecules to produce three CoA molecules as a side product and mevalonic
acid. Mevalonic acid is then Phosphorylated by ATP and decarboxylated to form
isopentyl pyrophosphate. Four of these molecules form geranylgeranyl
pyrophosphate which serves as the donor for all GA carbon atoms.
This compound is then converted to copalylpyrophos-phate
which has 2 ring systems. Copalylpyrophosphate is then converted to kaurene
which has 4-ring systems. Sub-sequent oxidations reveal kaurenol (alcohol
form), kaurenal (aldehyde form), and kaurenoic acid respectively.
Kaurenoic acid is converted to the aldehyde form of GA12 by
decarboxylation. GA12 is the first true gibberellane ring system with 20
carbons. From the aldehyde form of GA12 arise both 20 and 19 carbon
gibberellins but there are many mechanisms by which these other compounds
arise. During active growth, the plant will metabolize most gibberellins by
hydroxylation to inactive conjugates quickly with, the exception of GA3. GA3 is
degraded much slower which helps to explain why the symptoms initially
associ-ated with the hormone in the disease bakanae are present. Inactive
conjugates might be stored or translocated via the phloem and xylem before
their release (activation) at the proper time and in the proper tissue.
Functions
of gibberellins
· Stimulates stem elongation by
stimulating cell division and elongation. GA controls internode elongation in
the mature regions of plants. Dwarf plants do not make enough active forms of
GA.
· Flowering in biennial plants is
controlled by GA. Bien-nials grow one year as a rosette and after the winter,
they bolt (rapid expansion of internodes and formation of flowers).
· Breaks seed dormancy in some plants
that require strati-fication or light to induce germination.
· Stimulates α-amylase production in germinating cereal grains for
mobilization of seed reserves.
· Juvenility refers to the different
stages that plants may exist in. GA may help determine whether a particular
plant part is juvenile or adult.
· Stimulates germination of pollen and
growth of pollen tubes.
· Induces maleness in dioecious
flowers (sex expres-sion).
· Can cause parthenocarpic (seedless)
fruit development or increase the size of seedless fruit (grapes).
· Can delay senescence in leaves and
citrus fruits. May be involved in phytochrome responses.
Abscisic Acid
Natural growth inhibiting substances are present in plants
and affect the normal physiological process of them. One such compound is
abscisic acid, a single compound unlike the auxins, gibberellins, and
cytokinins. It was called ‘absci-sin II’ originally because it was thought to
play a major role in abscission of fruits. At about the same time another group
was calling it ‘dormin’ because they thought it had a major role in bud
dormancy. Though abscisic acid gener-ally is thought to play mostly inhibitory
roles, it has many promoting functions as well.
In 1963, when Frederick Addicott and his associates were the
one to identify abscisic acid. Two compounds were isolated and named as abscisin
I and abscisin II. Abscisin II is presently called abscisic acid (ABA). At the
same time Philip Wareing, who was studying bud dormancy in woody plants and Van
Steveninck, who was studying abscission of flowers and fruits discovered the
same compound.
Production
and occurrence
ABA is a naturally occurring sesquiterpenoid (15-carbon)
compound in plants, which is partially produced via the mevalonic pathway in
chloroplasts and other plastids. Because it is synthesized partially in the
chloroplasts, it makes sense that biosynthesis primarily occurs in the leaves.
The production of ABA is by stresses such as water loss and freezing
temperatures. The biosynthesis occurs indi-rectly through the production of
carotenoids. Breakdown of these carotenoids occurs by the following mechanism: Violaxanthin
(forty carbons) is isomerized and then splitted via an isomerase reaction
followed by an oxidation reaction. One molecule of xanthonin is produced from
one molecule of violaxanthonin and it is not clear what happens to the
remaining byproducts. The one molecule of xanthonin produced is unstable and
spontaneously changed to ABA aldehyde. Further oxidation results in ABA.
Activation of the molecule can occur by two methods. In the first, method, an
ABA-glucose ester can form by attachment of glucose to ABA. In the second
method, oxidation of ABA can occur to form phaseic acid and dihyhdrophaseic
acid. Both xylem and phloem tissues carries ABA. It can also be translocated
through parenchyma cells. Unlike auxins, ABA is capable of moving both up and
down the stem.
Functions
of abscisic acid
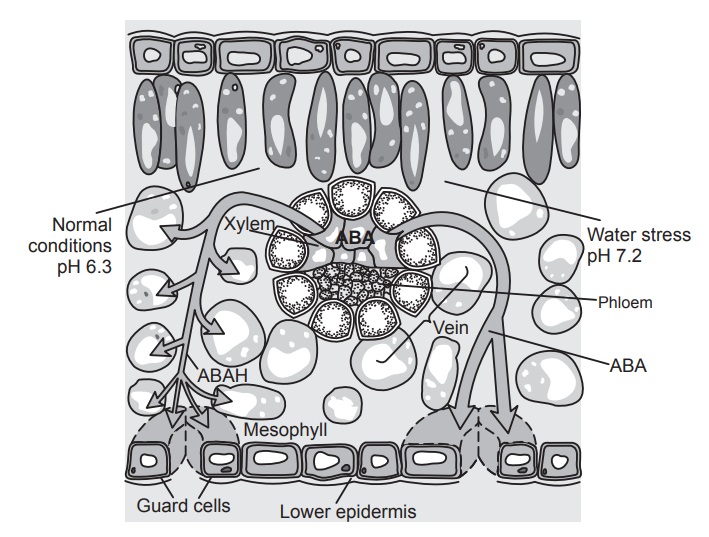
· The abscisic acid stimulates the
closure of stomata (water stress brings about an increase in ABA synthesis)
(Figure 6.3).
· Involved in abscission of buds,
leaves, petals, flowers, and fruits in many, if not all, instances, as well as
in dehiscence of fruits.
· Production is accentuated by
stresses such as water loss and freezing temperatures.
· Involved in bud dormancy.
· Prolongs seed dormancy and delays
germination (vivipary).
· Inhibits elongation.
· ABA is implicated in the control of
elongation, lateral root development, and geotropism, as well as in water
uptake and ion transport by roots.
· ABA coming from the plastids
promotes the metabolism of ripening.
· Promotes senescence.
· Can reverse the effects of growth
stimulating hormones.
Polyamines
Polyamines are unique as they are effective in relatively
high concentrations. Typical concentrations range from 5 to 500 mg/L.
Polyamines influence flowering and promote plant regeneration. Few examples are
Spermine, Spermidine and Putrescine. They play a major role in basic genetic
processes such as DNA synthesis and gene expression. Spermine and spermidine
bind to the phosphate backbone of nucleic acids. The interaction is mostly
based on electrostatic interactions between negatively charged phosphates of
the nucleic acids and the positively charged ammonium groups of the polyamines.
Polyamines are responsible for cell migration,
proliferation and differentiation in plants. They represent a group of plant
growth hormones, but they also have an effect on skin, hair growth, female
fertility, fat depots, pancreatic integrity and regenerative growth in mammals.
In addition, spermine is an important reagent widely used to precipitate DNA
in molecular biology protocols. Spermidine is a standard reagent in PCR
applications.
Spermine and spermidine are derivatives of putrescine
(1,4-diaminobutane) which is produced from L-ornithine by action of ODC
(ornithine decarboxylase). L-ornithine is the product of L-arginine degradation
by arginase. Spermidine is a triamine structure that is produced by
spermidine synthase (SpdS) which catalyses monoalkylation of putrescine
(1,4-diaminobutane) with decarboxylated S-adenosylmethionine (dcAdoMet)
3-aminopropyl donor. The formal alkylation of both amino groups of putrescine
with the 3-aminopropyl donor yields the symmetrical tetraamine spermine.
Brassinosteroids
There are approximately 60 naturally occurring polyhydroxy
steroids known as brassinosteroids (BRs). They are named after the first one
identified, brassinolide, which was isolated from rape in 1979. They appear to
be widely distributed in the plant kingdom.
In the early 1980s USDA scientists showed that BR could
increase yields of radishes, lettuce, beans, peppers and potatoes. However,
subsequent results under field conditions were disappointing because
inconsistent results were obtained. For this reason testing was phased out in
the United States. More recently large-scale field trials in China and Japan
over a six-year period have shown that 24-epibrassinolide, an alternative to
brassinolide, increased the production of agronomic and horticultural crops
(including wheat, corn, tobacco, watermelon, and cucumber). However, once again
depending on cultural conditions, method of application, and other factors, the
results sometimes were striking while other times they were marginal. Further
improvements in the formulation, application method, timing, effects of
environmental conditions, and other factors need to be investigated further in
order to identify the reason for these variable results.
Brassinosteroids may be a new class of plant growth
substances. They are widely distributed within the plant kingdom, they have an
effect at extremely low concentrations, both in bioassays and whole plants,
and they have a range of effects that are different from the other classes of
plant substances. Finally, they can be applied to one part of the plant and
transported to another location where, in very low amounts, they elicit a
biological response.
Functions
of brassinosteroids
· Promote shoot elongation at low
concentrations.
· Strongly inhibit root growth and development.
Promote ethylene biosynthesis and epinasty.
· Interfere with ecdysteroids
(moulting hormones) in insects.
· Have had contradictory effects in
tissue culture. 24-epi-brassinolide has been shown to mimic culture
condition-ing factors and to be synergistic with these factors in promoting
carrot cell growth. However, in transformed tobacco cells brassinosteroids in
low concentrations significantly inhibited cell growth.
· Enhance xylem differentiation.
· Decrease fruit abortion and drop.
· Enhance resistance to chilling,
disease, herbicide, and salt stress.
· Promotion of germination.
· Promote changes in plasmalemma
energization and transport, assimilate uptake.
· Increase RNA and DNA polymerase
activities and synthesis of RNA, DNA, and protein.
Salicylic Acid
Salicylic acid has been known to be present in some plant
tissues for quite some time, but has only recently been recognized as a
potential PGR. Salicylic acid is synthesized from the amino acid
phenylalanine. SA is thought by some to be a new class of plant growth
regulator. It is a chemically characterized compound, ubiquitously found in the
plant kingdom and has an effect on many physiological processes in plants at
low concentrations. Further molecular studies on SA signal transduction should
yield insights into the mechanism of action of this important regulatory
compound.
Functions
of salicylic acid
· Promotes flowering.
· Stimulates thermogenesis in Arum flowers.
· Stimulates plant pathogenesis
protein production (systemic acquired resistance).
· May enhance longevity of flowers.
· May inhibit ethylene biosynthesis.
· May inhibit seed germination.
Blocks the wound response. Reverses
the effects of ABA.
Jasmonates
Jasmonates are represented by
jasmonic acid and its methyl ester. They were first isolated from the jasmine
plant in which the methyl ester is an important product in the perfume
industry. Jasmonic acid is synthesized from lino-lenic acid, which is an
important fatty acid. Jasmonic acid is considered by some to be a new class of
plant growth regulator. It is a chemically characterized compound and has been
identified in many plant species. It has physiological effects at very low
concentrations and indirect evidence suggests that it is transported throughout
the plant.
Functions of jasmonates
·
Inhibition
of many processes such as seedling longitu-dinal growth, root length growth,
mycorrhizial fungi growth, tissue culture growth, embryogenesis, seed
germination, pollen germination, flower bud formation, carotenoid biosynthesis,
chlorophyll formation, rubisco biosynthesis, and photosynthetic activities
·
Promotion
of senescence, abscission, tuber formation, fruit ripening, pigment formation,
tendril coiling, differentiation in plant tissue culture, adventitious root
formation, breaking of seed dormancy, pollen germination, stomatal closure,
microtubule disruption, chlorophyll degradation, respiration, ethylene
biosynthesis, and protein synthesis
·
They
play an important role in plant defense by inducing proteinase synthesis.
Related Topics
