Cellular Components - Bacterial Ultrastructure
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Bacteria
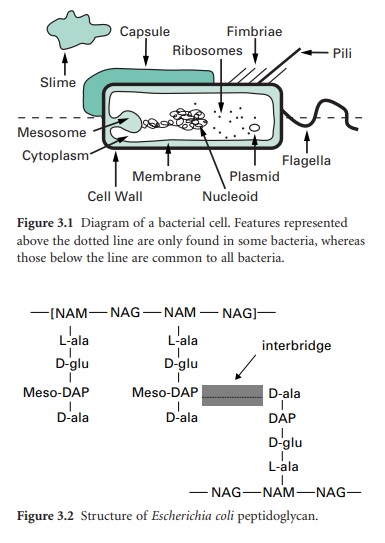
Compared with eukaryotic cells, bacteria possess a fairly simple base cell structure, comprising cell wall, cytoplasmic membrane, nucleoid, ribosomes and occasionally inclusion granules
CELLULAR COMPONENTS
Compared with eukaryotic
cells, bacteria possess a fairly simple base cell structure, comprising cell
wall, cytoplasmic membrane, nucleoid, ribosomes and occasionally inclusion
granules (Figure 3.1). Nevertheless it is important for several reasons to have
a good knowledge of these structures and their functions. First, the study of
bacteria provides an excellent route for probing the nature of biological
processes, many of which are shared by multicellular organisms. Secondly, at an
applied level, normal bacterial processes can be customized to benefit society
on a mass scale. Here, an obvious example is the largescale industrial
production (fermentation) of antibiotics. Thirdly, from a pharmaceutical and
healthcare perspective, it is important to be able to know how to kill bacterial
contaminants and disease-causing organisms. To treat infections antimicrobial
agents are used to inhibit the growth of bacteria, a process known as
antimicrobial chemotherapy. The essence of antimicrobial chemotherapy is
selective toxicity, which is achieved by exploiting differences between the
structure and metabolism of bacteria and host cells. Selective toxicity is,
therefore, most efficient when a similar target does not exist in the host.
Examples of such targets will be noted in the following sections.
Cell wall
The bacterial cell wall
is an extremely important structure, being essential for the maintenance of the
shape and integrity of the bacterial cell. It is also chemically unlike any
structure present in eukaryotic cells and is therefore an obvious target for
antibiotics that can attack and kill bacteria without harm to the host.
The primary function of
the cell wall is to provide a strong, rigid structural component that can
withstand the osmotic pressures caused by high chemical concentrations of
inorganic ions in the cell. Most bacterial cell walls have in common a unique structural
component called peptidoglycan (also called murein or glycopeptide); exceptions
include the mycoplasmas, extreme halophiles and the archaea. Peptidoglycan is a
large macromolecule containing glycan (polysaccharide) chains that are crosslinked
by short peptide bridges. The glycan chain acts as a backbone to peptidoglycan,
and is composed of alternating residues of N-acetyl
muramic acid (NAM) and N-acetyl
glucosamine (NAG). To each molecule of NAM is attached a tetrapeptide
consisting of the amino acids L-alanine, D-alanine, D-glutamic acid and either
lysine or diaminopimelic acid (DAP). This glycan tetrapeptide repeat unit is
crosslinked to adjacent glycan chains, either through a direct peptide linkage
or a peptide inter-bridge (Figure 3.2). The types and numbers of crosslinking
amino acids vary from organism to organism. Other unusual features of the cell
wall that provide potential antimicrobial targets are DAP and the presence of
two amino acids that have the D-configuration.
Bacteria can be divided
into two large groups, Gram-positive and Gram-negative, on the basis of a
differential staining technique called the Gram stain. Essentially, the Gram
stain consists of treating a film of bacteria dried on a microscope slide with
a solution of crystal violet, followed by a solution of iodine; these are then
washed with an alcohol solution. In Gram negative organisms the cells lose the
crystal violet–iodine complex and are rendered colourless, whereas Gram-positive
cells retain the dye. Regardless, both cell types are counterstained with a different
coloured dye, e.g. carbolfuchsin, which is red.
Hence, under the light
microscope Gram-negative cells appear red while Gram-positive cells are purple.
These marked differences in response reflect differences in cell wall
structure. The Gram-positive cell wall consists primarily of a single type of
molecule whereas the Gram-negative cell wall is a multi-layered structure and quite
complex.
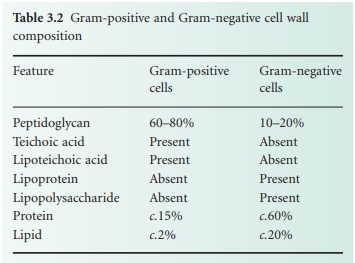
The cell walls of Gram-positive
bacteria are quite thick (20–80 nm) and consist of between 60% and 80% peptidoglycan,
which is extensively crosslinked in three dimensions to form a thick polymeric
mesh (Figure 3.3). Gram-positive walls frequently contain acidic polysaccharides
called teichoic acids; these are either ribitol phosphate or glycerol phosphate
molecules that are connected by phosphodiester bridges. Because they are
negatively charged, teichoic acids are partially responsible for the negative
charge of the cell surface as a whole.
Their function may be to
effect passage of metal cations through the cell wall. In some Gram-positive
bacteria glycerol–teichoic acids are bound to membrane lipids and are termed
lipoteichoic acids. During an infection, lipoteichoic acid molecules released
by killed bacteria trigger an inflammatory response. Cell wall proteins, if
present, are generally found on the outer surface of the peptidoglycan.
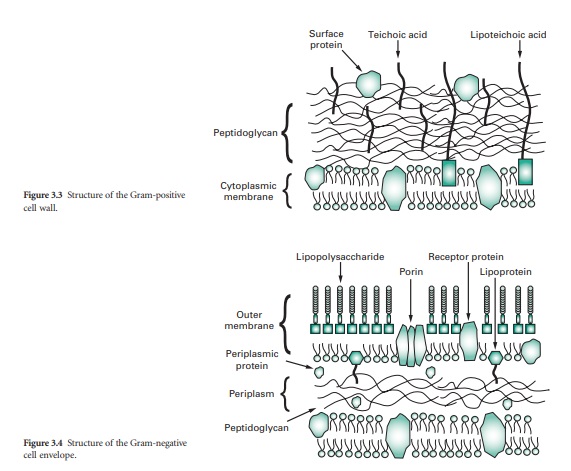
The wall, or more
correctly, envelope of Gram-negative cells is a far more complicated structure
(Figure 3.4). Although it contains less peptidoglycan (10–20% of wall), a
second membrane structure is found outside the peptidoglycan layer. This outer
membrane is asymmetrical, composed of proteins, lipoproteins, phospholipids and
a component unique to Gram-negative bacteria, lipopolysaccharide (LPS). Essentially,
the outer membrane is attached to the peptidoglycan by a lipoprotein, one end
of which is covalently attached to peptidoglycan and the other end is embedded
in the outer membrane. The outer membrane is not a phospholipid bilayer
although it does contain phospholipids in the inner leaf, and its outer layer
is composed of LPS, a polysaccharide–lipid molecule. Proteins are also found in
the outer membrane, some of which form trimers that traverse the whole membrane
and in so doing form waterfilled channels or porins through which small molecules
can pass. Other proteins are found at either the inner or outer face of the
membrane.
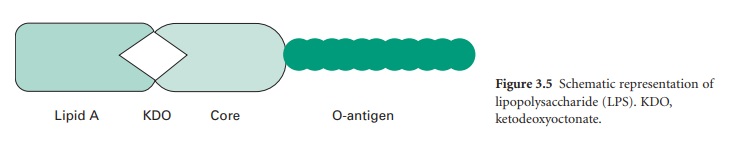
The LPS (Figure 3.5) is
an important molecule because it determines the antigenicity of the Gram-negative
cell and it is extremely toxic to animal cells. The molecule consists of three
regions, namely lipid A, core polysaccharide and Ospecific polysaccharide. The
lipid A portion is composed of a disaccharide of glucosamine phosphate bound to
fatty acids and forms the outer leaflet of the membrane. It is the lipid A
component that is responsible for the toxic and pyrogenic properties of Gram-negative
bacteria. Lipid A is linked to the core polysaccharide by the unique molecule
ketodeoxyoctonate (KDO), and at the other end of the core is the Opolysaccharide
(Oantigen), which usually contains sixcarbon sugars as well as one or more
unusual deoxy sugars such as abequose.
Although the outer
membrane is relatively permeable to small molecules, it is not permeable to
enzymes or large molecules. Indeed, one of the major functions of the outer
membrane may be to keep certain enzymes that are present outside the
cytoplasmic membrane from diffusing away from the cell. Moreover, the outer
membrane is not readily penetrated by hydrophobic compounds and is, therefore,
resistant to dissolution by detergents.
The region between the
outer surface of the cytoplasmic membrane and the inner surface of the outer
membrane is called the periplasm. This occupies a distance of about 12–15 nm,
is gel like in consistency and, in addition to the peptidoglycan, contains
sugars and an abundance of proteins including hydrolytic enzymes and transport
proteins. Table 3.2 summarizes the major differences in wall composition
between Gram-positive and Gram-negative cells.
Cytoplasmic membrane
Biochemically, the
cytoplasmic membrane is a fragile, phospholipid bilayer with proteins
distributed randomly throughout. These are involved in the various transport
and enzyme functions associated with the membrane. A major difference in
chemical composition between prokaryotic and eukaryotic cells is that
eukaryotes have sterols in their membranes (e.g. cholesterol) whereas
prokaryotes do not. The cytoplasmic membrane serves many functions, including transport
of nutrients, energy generation and electron transport; it is the location for
regulatory proteins and biosynthetic proteins, and it acts as a semipermeable
selectivity barrier between the cytoplasm and the cell environment.
Invaginations of the cytoplasmic
membrane are referred to as mesosomes.
Those that form near the septum of Gram-positive cells serve as organs of
attachment for the bacterial chromosome.
Pppppppppppppppppppppppppppppppp
Cytoplasm
The cytoplasm consists
of approximately 80% water and contains enzymes that generate ATP directly by
oxidizing glucose and other carbon sources. The cytoplasm also contains some of
the enzymes involved in the synthesis of peptidoglycan subunits. Ribosomes, the
DNA genome (nucleoid) and inclusion granules are also found in the cytoplasm.
Nucleoid
The bacterial chromosome
exists as a singular, covalently closed circular molecule of double-stranded
DNA comprising approximately 4600 kilobase pairs. It is complexed with small
amounts of proteins and RNA, but unlike eukaryotic DNA, is not associated with
histones. The DNA, if linearized, would be about 1 mm in length. In order to
package this amount of material the cell requires that the DNA is supercoiled
into a number of domains (c.50) and
that the domains are associated with each other and stabilized by specific proteins
into an aggregated mass or nucleoid. The enzymes, topoisomerases, that control
topological changes in DNA architecture are different from their eukaryotic
counterparts (which act on linear chromosomes) and therefore provide a unique
biochemical target for antibiotic action.
Plasmids
Plasmids are relatively
small, circular pieces of double-stranded extrachromosomal DNA. They are
capable of autonomous replication and encode for many auxiliary functions that
are not usually necessary for bacterial growth. One such function of great
significance is that of antibiotic resistance (Chapter 13). Plasmids may also
transfer readily from one organism to another, and between species, thereby
increasing the spread of resistance.
Ribosomes
The cytoplasm is densely
packed with ribosomes. Unlike eukaryotic cells these are not associated with a
membranous structure; the endoplasmic reticulum is not a component of
prokaryotic cells. Bacterial ribosomes are 70S in size, made up of two subunits
of 30S and 50S. This is smaller than eukaryotic ribosomes, which are 80S in
size (40S and 60S subunits). Differences will therefore exist in the size and
geometry of RNA binding sites.
Inclusion granules
Bacteria occasionally
contain inclusion granules within their cytoplasm. These consist of storage
material composed of carbon, nitrogen, sulphur or phosphorus and are formed
when these materials are replete in the environment to act as repositories of these
nutrients when shortages occur. Examples include polyβhydroxybutyrate, glycogen and polyphosphate.
Cell surface components
The surface of the
bacterial cell is the portion of the organism that interacts with the external
environment most directly. As a consequence, many bacteria deploy components on
their surfaces in a variety of ways that allow them to withstand and survive
fluctuations in the growth environment. The following sections describe a few
of these components that are commonly found, although not universally, that
allow bacteria to move, sense their environment, attach to surfaces and provide
protection from harsh conditions.
Flagella
Bacterial motility is
commonly provided by flagella, long (c.12
μm) helical-shaped structures that project from
the surface of the cell. The filament of the flagellum is built up from
multiple copies of the protein flagellin. Where the filament enters the surface
of the bacterium, there is a hook in the flagellum, which is attached to the
cell surface by a series of complex proteins called the flagellar motor. This
rotates the flagellum, causing the bacterium to move through the environment.
The numbers and distribution of flagella vary with bacterial species. Some have
a single, polar flagellum, whereas others are flagellate over their entire
surface (peritrichous); intermediate forms also exist.
Fimbriae
Fimbriae are
structurally similar to flagella, but are not involved in motility. Although
they are straighter, more numerous and considerably thinner and shorter (3 μm) than flagella, they do consist of protein and
project from the cell surface. There is strong evidence to suggest that
fimbriae act primarily as adhesins, allowing organisms to attach to surfaces,
including animal tissues in the case of some pathogenic bacteria, and to initiate
biofilm formation. Fimbriae are also responsible for haem-agglutination and
cell clumping in bacteria. Among the best characterized fimbriae are the type I
fimbriae of enteric (intestinal) bacteria.
Pili
Pili are morphologically
and chemically similar to fimbriae, but they are present in much smaller
numbers (<10) and are usually
longer. They are involved in the genetic exchange process of conjugation.
Capsules and slime layers
Many bacteria secrete
extracellular polysaccharides (EPS) that are associated with the exterior of
the bacterial cell. The EPS is composed primarily of c.2% carbohydrate and 98% water, and provides a gummy exterior to the
cell. Morphologically, two extreme forms exist: capsules, which form a tight, fairly rigid layer closely associated
with the cell, and slimes , which are
loosely associated with the cell. Both forms function similarly, to offer
protection against desiccation, to provide a protective barrier against the
penetration of biocides, disinfectants and positively charged antibiotics, to
protect against engulfment by phagocytes and protozoa and to act as a cement
binding cells to each other and to the substratum in biofilms (see below). One
such polymer that performs all these functions is alginate, produced by Pseudomonas aeruginosa; dextran,
produced by Leuconostoc mesenteroides,
is another. Both polymers may be harvested and used variously as pharmaceutical
aids, surgical dressings and drug delivery systems, although the preferred
source of alginate is seaweed rather than bacteria.
S-layers
S-layers are the most
common cell wall type amongst the archaea. These consist of a two-dimensional
para-crystalline array of proteins or glycoproteins which show various ordered
symmetries when viewed under the electron microscope. In many species of
bacteria, Slayers are present on their outer surfaces in addition to other cell
wall components such as polysaccharides. In such arrangements the Slayer is
always the outermost layer. In addition to increasing the structural robustness
of the cell, S layers can act to a
certain extent as an external permeability barrier.
Related Topics
