Clonal Selection and Expansion
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Immunology
Within the body there may exist at any one time only a handful of naive B-lymphocytes capable of recognizing the same epitope.
CLONAL SELECTION AND EXPANSION
Within the body there may exist at any one
time only a handful of naive B-lymphocytes capable of recognizing the same
epitope. The meeting of an antigen and a naive B-lymphocyte capable of
recognizing an epitope within the antigen occurs through the delivery of
antigen to lymphoid tissues of the spleen, lymph nodes and local lymphoid
tissue within mucosal surfaces (mucosal associated lymphoid tissue, MALT) and
skin (SALT). This lymphoid tissue is rich in lymphocytes. Further, a proportion
of B-lymphocytes will always be recirculating from the lymphoid tissue through
the lymph and blood circulations and so able to encounter circulating antigen.
Antigen will be specifically recognized by IgM molecules present on the surface
of the naive B-lymphocyte. Following this antigen-driven selection of a
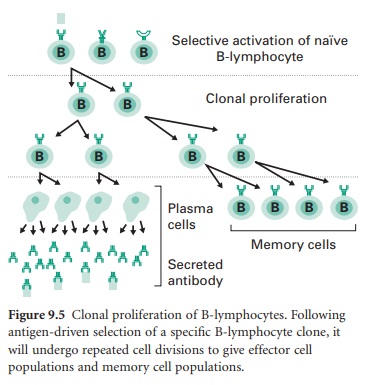
specific B-lymphocyte clone, the clone will undergo repeated cell divisions.
Some of the daughter cells will differentiate into short-lived (2–3 days)
plasma cells able to secrete antibody of different classes to combat the initial
primary antigen exposure. Other clonal daughter cells will become long-lived
B-lymphocyte memory cells populating the lymphoid tissue and spreading around
the body through the lymph and blood circulations. These cells will provide
‘immunological memory’ able to generate a more rapid and pronounced secondary
response on subsequent exposure to the original antigen (Figure 9.5).
This process of clonal selection and
expansion to form memory cell populations is the basis of vaccination. The
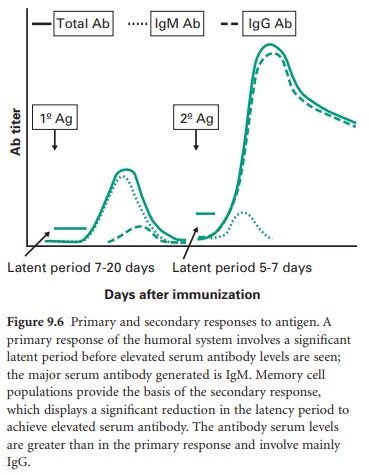
initial introduction of antigen gives rise to a primary response (Figure 9.6)
in which there is a significant latent period before increased serum antibody
levels are observed; the main antibody response is IgM production, although
some IgG is also synthesized and secreted. On re-exposure to the same antigen a
secondary response is elicited. The features of the secondary response include:
• a
reduced latent period between antigen challenge and increases in serum antibody
(e.g. latent period of 5–7 days for the secondary response vs 7–20 days for the
primary response)
• an
antibody response dominated by IgG which is more pronounced with higher serum
levels achieved.
In the absence of helper T-lymphocyte
involvement (T-cell-independent humoral responses, e.g. where antigen is
carbohydrate alone) B-lymphocyte memory cell populations are not produced, and
antibody class switching is restricted. Hence, under these circumstances, the
primary and secondary antibody responses to antigen challenge are essentially
indistinguishable and exhibit a prolonged latent period, relatively low levels
of serum antibody produced, and IgM as the main serum antibody.
Related Topics
