The Building Blocks
| Home | | Pharmacognosy |Chapter: Pharmacognosy and Phytochemistry : General Biosynthetic Pathways of Secondary Metabolites
The building blocks for secondary metabolites are derived from primary metabolism as indicated in Figure below. This scheme outlines how metabolites from the fundamental processes of photosynthesis, glycolysis, and the Krebs cycle are tapped off from energy-generating processes to provide biosynthetic intermediates.
THE BUILDING BLOCKS
The building blocks for secondary metabolites are derived from primary metabolism as indicated in Figure below. This scheme outlines how metabolites from the fundamental processes of photosynthesis, glycolysis, and the Krebs cycle are tapped off from energy-generating processes to provide biosynthetic intermediates. The number of building blocks needed is surprisingly few, and as with any child’s construction set a vast array of objects can be built up from a limited number of basic building blocks. By far the most important building blocks employed in the biosynthesis of secondary metabolites are derived from the intermediates acetyl coenzyme A (acetyl-CoA), shikimic acid, mevalonic acid, and 1-deoxyxylulose 5-phosphate. These are utilized respectively in the acetate, shikimate, mevalonate, and deoxyxylulose phosphate pathways.
In addition to acetyl-CoA, shikimic acid, mevalonic acid, and deoxyxylulose phosphate, other building blocks based on amino acids are frequently employed in natural product synthesis.
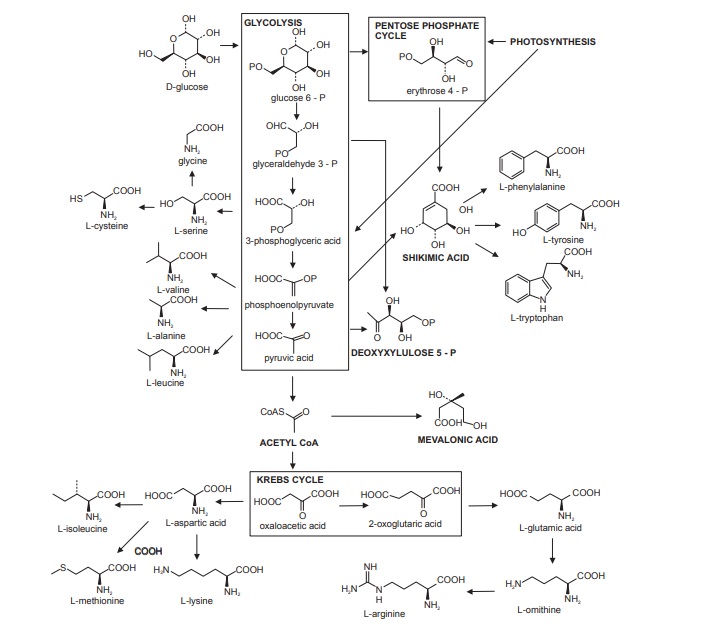
Peptides, proteins, alkaloids, and many antibiotics are derived from amino acids, and the origins of the most important amino acid components of these are briefly indicated in Figure above. Intermediates from the glycolytic pathway and the Krebs cycle are used in constructing many of them, but the aromatic amino acids phenylalanine, tyrosine, and tryptophan are themselves products from the shikimate pathway. Ornithine, a nonprotein amino acid, and its homologue lysine, are important alkaloid precursors having their origins in Krebs cycle intermediates.
Relatively few building blocks are routinely employed, and the following list, though not comprehensive, includes those most frequently encountered in producing the carbon and nitrogen skeleton of a natural product.
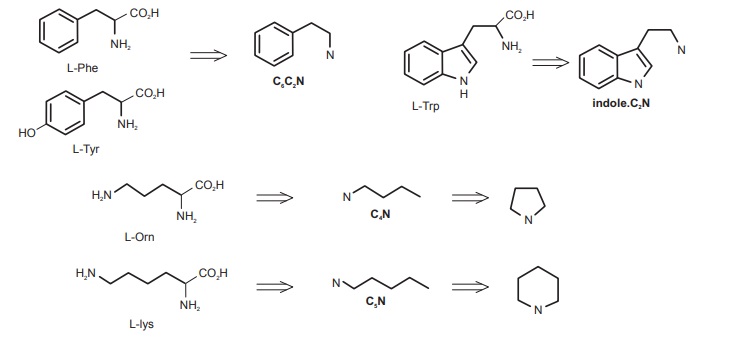
The structural features of these building blocks are shown in Figure below.
C1: The simplest of the building blocks is composed of a single carbon atom, usually in the form of a methyl group, and most frequently it is attached to oxygen or nitrogen, but occasionally to carbon. It is derived from the S-methyl of L-methionine. The methylenedioxy group (OCH2O) is also an example of a C1 unit.
C2: A two-carbon unit may be supplied by acetyl-CoA. This could be a simple acetyl group, as in an ester, but more frequently it forms part of a long alkyl chain (as in a fatty acid) or may be part of an aromatic system (e.g. phenols). Of particular relevance is that in the latter examples, acetyl-CoA is first converted into the more reactive malonyl-CoA before its incorporation.
C5: The branched-chain C5 ‘isoprene’ unit is a feature of compounds formed from mevalonate or deoxyxylulose phosphate. Mevalonate itself is the product from three acetyl-CoA molecules, but only five of mevalonate’s six carbons are used, the carboxyl group being lost. The alternative precursor deoxyxylulose phosphate, a straight-chain sugar derivative, undergoes a skeletal rearrangement to form the branched chain isoprene unit.
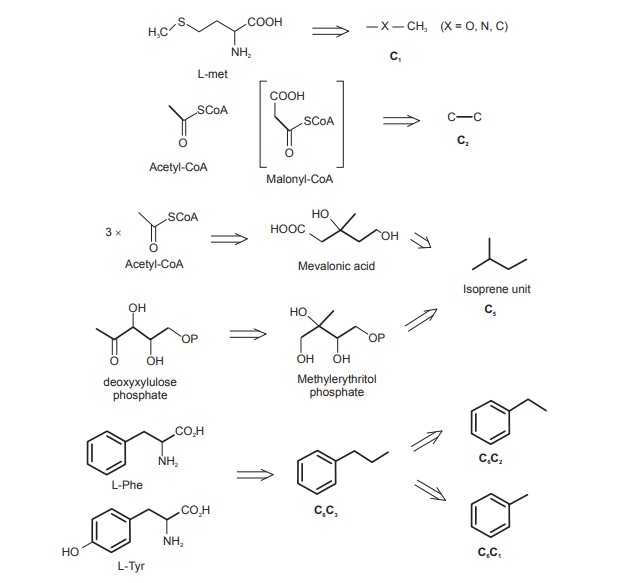
C6C3: This refers to a phenylpropyl unit and is obtained from the carbon skeleton of either L-phenylalanine or L-tyrosine, two of the shikimate-derived aromatic amino acids. This, of course, requires loss of the amino group. The C3 side chain may be saturated or unsaturated, and may be oxygenated.
Sometimes the side chain is cleaved, removing one or two carbons. Thus, C6C2 and C6C1 units represent modified shortened forms of the C6C3system.
C6C2N: Again, this building block is formed from either L-phenylalanine or L-tyrosine, L-tyrosine being by far the more common. In the elaboration of this unit, the carboxyl carbon of the amino acid is removed.
Indole.C2N: The third of the aromatic amino acids is L-tryptophan. This indole-containing system can undergo decarboxylation in a similar way to L-phenylalanine and L-tyrosine so providing the remainder of the skeleton as an indole. C2N unit.
C4N: The C4N unit is usually found as a heterocyclic pyrrolidine system and is produced from the nonprotein amino acid L-ornithine. In marked contrast to the C6C2N and indole.C2N units described above, ornithine supplies not its α-amino nitrogen, but the δ-amino nitrogen. The carboxylic acid function and the α-amino nitrogen are both lost.
C5N: This is produced in exactly the same way as the C4N unit, but using L-lysine as precursor. The ε-amino nitrogen is retained, and the unit tends to be found as a piperidine ring system.
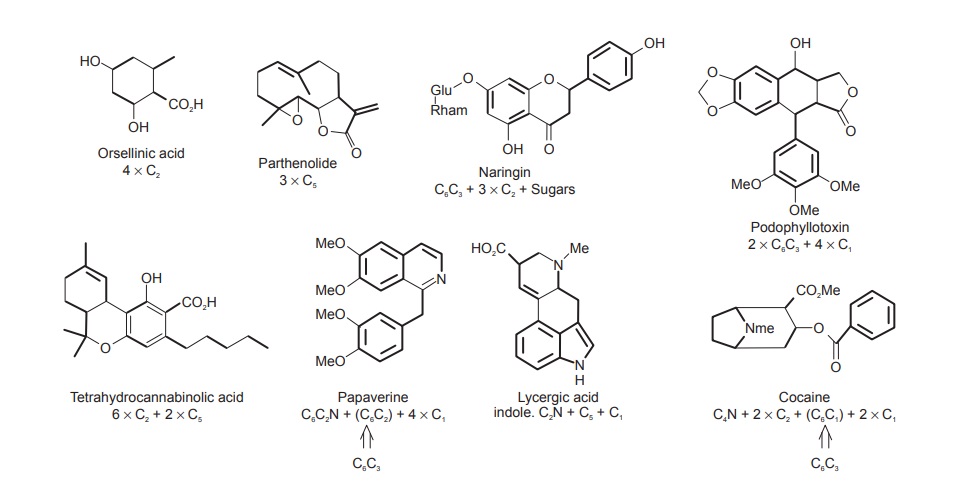
These eight building blocks form the basis of many of the natural product structures discussed in this chapter. Simple examples of how compounds can be visualized as a combination of building blocks are shown in Figure below.
Although primary and secondary metabolism are inter-related to the extent that an absolute distinction is meaningless, for the purpose of this chapter some division has had to be made, and this has been based on biosynthetic pathways. Excluding the primary processes of sugar and protein biosynthesis, there are three main routes to the wealth of chemical compounds found in plants, that is, shikimic acid pathways, acetate-malonate, and acetate-mevalonate pathways, which are interrelated as shown in figure below.
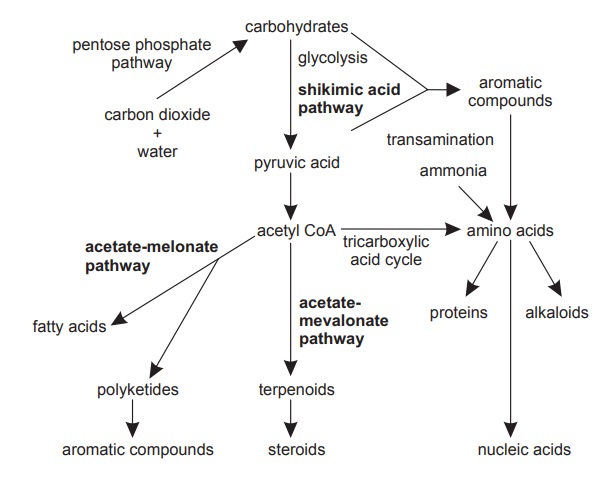
Biosynthetic pathways in plants
Shikimic Acid Pathway
The shikimate pathway provides an alternative route to aromatic compounds, particularly the aromatic amino acids L-phenylalanine, L-tyrosine, and L-tryptophan. This pathway is employed by microorganisms and plants, but not by animals, and accordingly the aromatic amino acids feature among those essential amino acids for men whom have to be obtained in the diet. A central intermediate in the pathway is shikimic acid, a compound which had been isolated from plants of Illicium species (Japanese ‘shikimi’) many years before its role in metabolism had been discovered. Most of the intermediates in the pathway were identified by a careful study of a series of Escherichia coli mutants prepared by UV irradiation. Their nutritional requirements for growth, and any by-products formed, were then characterized. A mutant strain capable of growth usually differs from its parent in only a single gene, and the usual effect is the impaired synthesis of a single enzyme. Typically, a mutant blocked in the transformation of compound A into compound B will require B for growth whilst accumulating A in its culture medium. In this way, the pathway from phosphoenolpyruvate (from glycolysis) and D-rythrose 4-phosphate (from the pentose phosphate cycle) to the aromatic amino acids was broadly outlined. Phenylalanine and tyrosine form the basis of C6C3 phenylpropane units found in many natural products, for example, cinnamic acids, coumarins, lignans, and flavonoids, and along with tryptophan are precursors of a wide range of alkaloid structures. In addition, it is found that many simple benzoic acid derivatives, for example, gallic acid and p-aminobenzoic acid (4-aminobenzoic acid) are produced via branchpoints in the shikimate pathway (Figure below).
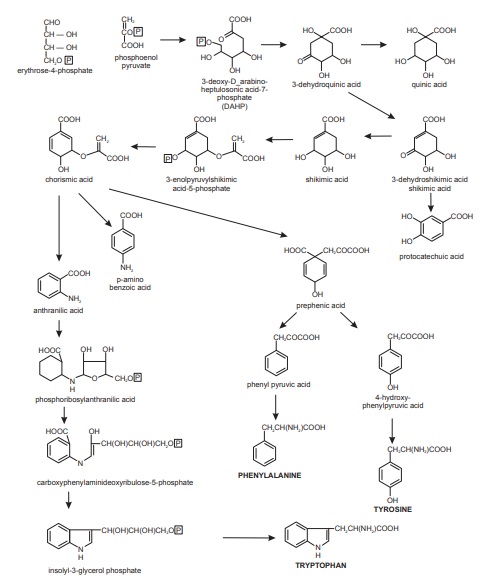
Shikimic acid pathway
The precursors, that is, D-erythrose 4-phosphate and phosphoenolpyruvate combine to form 3-deoxy-D-arab-ino-heptulosonic acid-7-phosphate (DAHP), a reaction catalysed by phospho-2-oxo-3-deoxyheptonate aldolase. The enzyme, 3-dehydroquinate synthase, catalysing the cyclization of DAHP to 3-dehydroquinic acid, requires cobalt (II) and nicotinamide adenine dinucleotide (NAD) as cofactors.
The shikimic acid pathway contains several branch points, the first of these, dehydroquinic acid, can be converted either to 3-dehydroshikimic acid, which continues the pathway, or to quinic acid. The enzymes catalysing the dehydration of dehydroquinic acid are of two kinds. Form 1, associated with shikimate dehydrogenase, is independent of shikimate concentration, while form 2 is specifically activated by shikimate.
It has been suggested that the two forms provide a control in the utilization of dehydroquinic acid producing either shikimic acid or protocatechuic acid.
After phosphorylation, catalysed by shikimate kinase, shikimic acid adds on enol pyruvate to form 3-enolpyru-vylshikimic acid-5-phosphate. This reaction is catalysed by enolpyruvylshikimate phosphate synthase, whereas conversion to chorismic acid is catalysed by chorismate synthase.
The formation of chorismic acid is an important branch point in the shikimic acid pathway as this compound can undergo three different types of conversion. The name ‘chorismic’ is derived from a Greek word for separate, indicating the multiple role of this compound. In the presence of glutamine, chorismic acid is converted to anthranilic acid, whereas chorismate mutase catalyses the formation of prephenic acid. chorismic acid is also converted into p-aminobenzoic acid.
Then after anthranilic acid is converted first to phos-phoribosylanthranilic acid and then to carboxyphenylamin-odeoxyribulose-5-phosphate, these reactions being catalysed by anthranilate phosphoribosyl transferase and phosphori-bosylanthranilate isomerase, respectively. Ring closure to form indolyl-3-glycerol phosphate is catalysed by indolyl-glycerol phosphate synthase. The enzyme catalysing the final reaction, that is, tryptophan synthase consists of two components; component A catalyses the dissociation of indolylglycerol phosphate to indole and glyceraldehydes-3-phosphate, whereas component B catalyses the direct condensation of indole with serine to form tryptophan.
Tyrosine and phenylalanine are both biosynthesized from prephanic acid, but by independent pathways.
In the formation of tyrosine, prephanic acid is first aromatized to 4-hydroxyphenylpyruvic acid, a reaction catalysed by prephenate dehydrogenase. Transamination, catalysed by tyrosine aminotransferase, then gives tyrosine.
The biosynthesis of phenylalanine involves first the aromatization of prephanic acid to phenylpyruvic acid, a reaction catalysed by prephenate dehydratase, and then transamination catalysed by phenylalanine aminotransferase, which gives phenylalanine.
Acetate-Mevalonate Pathway
Since a long time biochemists were aware of the involvement of acetic acid in the synthesis of cholesterol, squalene and rubber-like compounds. The discovery of acetyl coenzyme A called as ‘active acetate’ in 1950, further supported the role of acetic acid in biogenetic pathways. Later, mevalonic acid was found to be associated with the acetate. Mevalonic acid further produced isopentenyl pyrophosphate (IPP) and its isomer dimethylallyl pyrophosphate (DMAPP). These two main intermediates IPP and DMAPP set the ‘active isoprene’ unit as a basic building block of isoprenoid com-pounds. Both of these units yield geranyl pyrophosphate (C10-monoterpenes) which further association with IPP produces farnesyl pyrophosphate (C15-sesquiterpenes).
Farnesyl pyrophosphate with one more unit of IPP devel-ops into geranyl—eranyl pyrophosphate (C20-diterpenes). The farnesyl pyrophosphate multiplies with its own unit to produce squalene, and its subsequent cyclization gives rise to cyclopentanoperhydrophenantherene skeleton containing steroidal compounds like cholesterols and other groups like triterpenoids. The acetate mevalonate pathway thus works through IPP and DMAPP via squalene to produce two different skeleton containing compounds, that is, steroids and triterpenoids. It also produces vast array of monoterpenoids, sesquiterpenoids, diterpenoids, carotenoids, polyprenols, and also the compounds like glycosides and alkaloids in association with other pathways (Figure below).
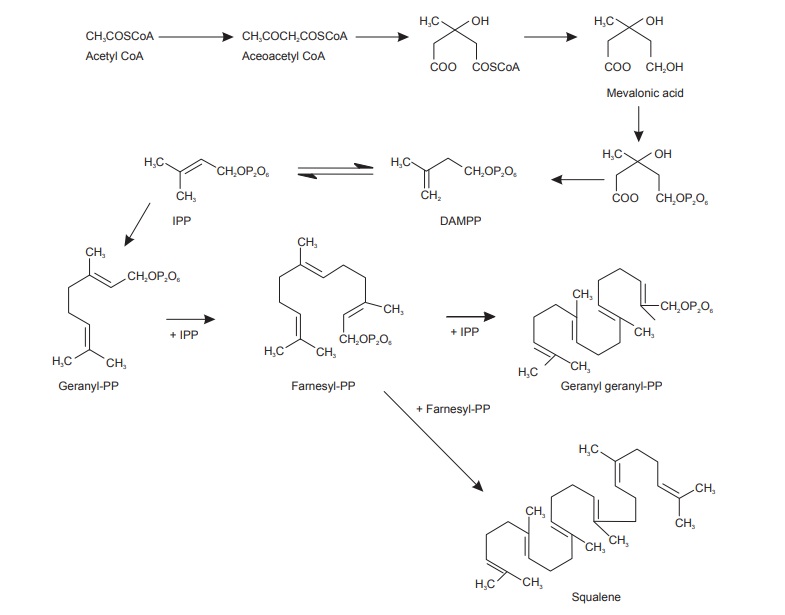
Acetate—mevalonate pathway
Acetate-Malonate Pathway
Acetate pathway operates functionally with the involvement of acyl carrier protein (ACP) to yield fatty acyl thioesters of ACP. These acyl thioesters forms the important intermediates in fatty acid synthesis. These C2 acetyl CoA units at the later stage produces even number of fatty acids from n-tetranoic (butyric) to n-ecosanoic (arachidic acid). The synthesis of fatty acids is thus explained by the reactions given in Figure below. Unsaturated fatty acids are produced by subsequent direct dehydrogenation of saturated fatty acids. Enzymes play important role in governing the position of newly introduced double bonds in the fatty acids.
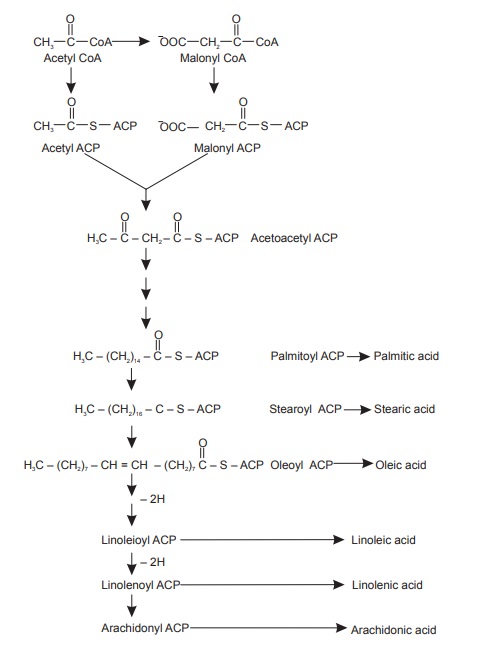
Acetate—malonate pathway
Related Topics
