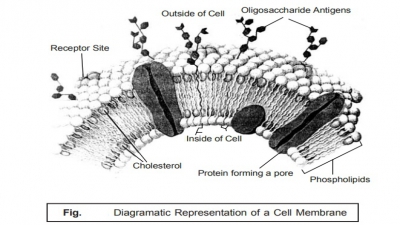
Archaeobacteria and Eubacteria
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Structure and Function of Bacterial Cells
The ‘archaeobacteria’ are unusual organisms by nature, and this particular category is known to comprise essentially of three different types of bacteria, namely : (1) Methanogenic bacteria, (2) Extreme halophiles, and (3) Thermoacidophiles
ARCHAEOBACTERIA
AND EUBACTERIA
It has
been observed that ‘all cells’
categorically fall into either of the two groups, namely : the eukaryotes, and the prokaryotes. Besides, the multicellular plants and animals are invariably eukaryotic in nature and character, and so are the numerous unicellular organisms. The only prokaryotes are the organisms, such as
: cyanobacteria (Gr. hyanos = dark blue). In the recent
past this very classification has undergone a considerable change. It has been
duly established and observed that there exists another ‘group of organisms’ amongst the bacteria that do not seem to fall
into either of the two aforesaid categories.
These organisms have been termed as the archaeobacteria,
which essentially designate an altogether new primary kingdom having an
entirely different status in the history and the natural order of life.
The
enormous volume of informations based on experimental evidences gathered from
studies of ribosomal RNA suggests
that archaeobacteria and eubacteria strategically got separated
at a very early stage in the pioneer process of evolution of life on this
planet (earth). Importantly, the phylogenetic*
distance that critically prevails between the two above mentioned categories of bacteria is reflected by some phenotypic** differences prominently,
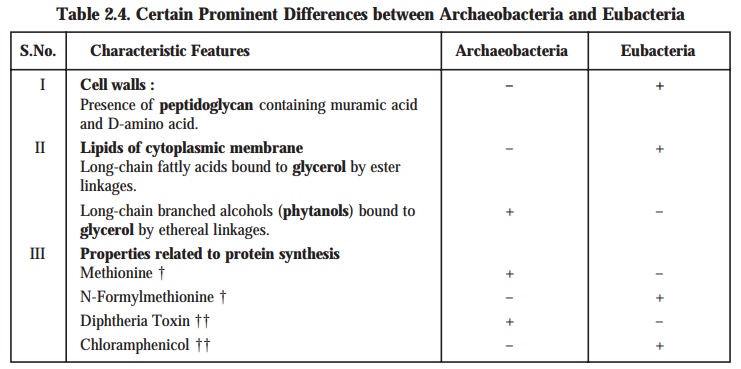
which may be summarized in the following Table : 2.4.
Archaeobacteria, in reality, do not represent a
perfect homogeneous group. One may, however, observe a substantial degree of heterogeneity amongst the eubacteria, so do the different types
of archaeobacteria specifically
differ from each other with respect to morphology, metabolism, chemical composition,
and habitat.
The ‘archaeobacteria’ are unusual organisms
by nature, and this particular category is known to comprise essentially of three different types of bacteria, namely :
(1) Methanogenic
bacteria,
(2) Extreme
halophiles, and
(3) Thermoacidophiles
These three groups of organisms shall now be
treated individually in the sections that follows :
1. Methanogenic Bacteria [Methanogens]
The methanogenic bacteria are considered to
be the hard-core anaerobes which,
invariably possess the capability of deriving energy for their progressive
growth by certain particular oxidizing chemical entities, for instance :
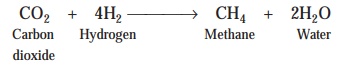
hydrogen (H2), formic acid (HCOOH) ; and actually exert their ‘action’ by making use of the electrons
thus produced to reduce ultimately carbon-dioxide (CO2) to give rise
to the formation of methane (CH4) gas :
It has
been observed that certain genera specifically may grow as autotrophs* — thereby utiliz-ing hydrogen and carbon dioxide as
exclusive sources of carbon as well as energy ; whereas some others do need
several additional components, for instance : organic-sulphur compounds, amino
acids, acetic acid, and vitamins. Interestingly, a plethora of species actually
grow quite abundantly and aggressively in a complex
media viz., comprising of yeast
extract in comparison to inorganic-salts
containing media.
Coenzymes** : There are at least two uncommon coenzymes that invariably
occur in all meth-anogenic bacteria
(methanogens) that have not been noticed in other varieties of
microorganisms.
Examples : Following are two typical examples of
methanogenic coenzymes :
(a) Coenzyme M — directly involved in methyl transfer reactions, and
(b) Coenzyme F420 — a flavin-like chemical entity
intimately involved in the anaerobic
elec-tron transport system of these microorganisms. It has the ability to fluoresce when exposed to UV light ; and, therefore, its presence may be detected by
visualizing the organisms via a fluorescence microscope conveniently
(also used for its critical identification and examina-tion).
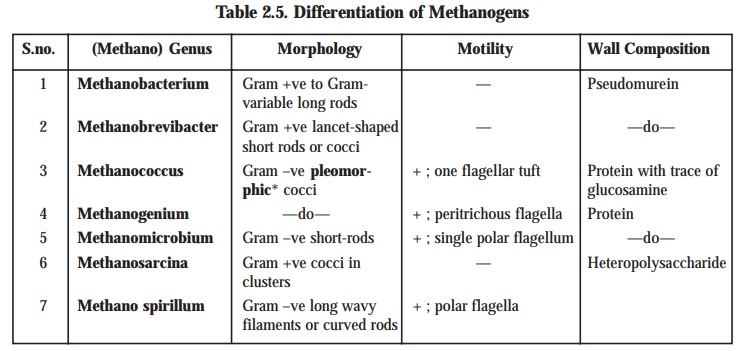
Differentiation of Methanogens : The
genera of methanogens i.e., the methane-producing bac-teria may be clearly differentiated
exclusively based upon their morphology***
and Gram reaction****. However, the
glaring distinct differences occurring in the cell-wall composition have been
duly observed to correlate specifically with these genera.
Table 2.5 : records the morphology, motility, and wall composition of several methanogenic organisms with specific ‘genus’.
Importantly,
the cell walls of two genera
essentially consist of pseudomurein,
that prominently differs from eubacterial peptidoglycan
by the following two distinct
structural features, namely :
(a) substitution
of N-acetyltalosaminuronic acid for N-acetylmuramic acid, and
(b) presence
of tetrapeptide composed totally of L-amino acids, having glutamic acid
attached duly at the C-terminal end.
Habitats : Interestingly, the methanogenic bacteria most commonly
found in anaerobic habi-tats that are eventually rich organic matter which
ultimately produced by nonmethanogenic
bacteria via fermentation to yield H2 and CO2. A few
such common as well as vital habitats are, namely : marine sediments, swamps,
marshes, pond and lake mud, intestinal tract of humans (GIT) and animals, rumen
of cattle (e.g., cow, buffalow,
sheep, pig, goat etc.), and anaerobic sludge digesters in sewage-treatment
plants.
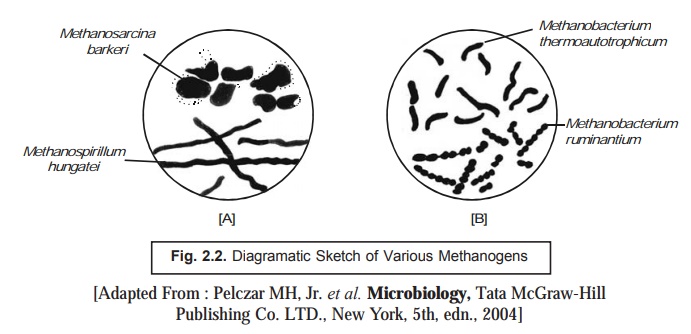
Figure.
2.6 [A and B] depicts the diagramatic sketch of the cells commonly observed in
various kinds of methanogenic organisms (viz.,
methane-producing bacteria).
Figure
2.6 [A] evidently shows the typical cells of Methanosarcina barkeri and Methanospirillum
hungatei representing ideally the
methane-producing bacteria.
Figure
2.6 [B] likewise illustrates the characteristic cells of Methanobacterium thermo-autotrophicum and Methanobacterium ruminantium designating the methanogens.
2. Extreme Halophiles
The extreme halophiles are aerobic
organisms and chemoorganotrophic* in nature that essen-tially need nearly 17 to
23% (w/v) sodium chloride (NaCl) for their normal and good growth. These extreme halophiles invariably stain
Gram-negative organisms that specifically vary from the rod or disk-shaped cells (i.e., the genus Halobacterium) to spherical or ovoid cocci (i.e., the genus Halococcus).
Habitat : They are most commonly found in ‘salt lakes’, such as :
·
The Great Salt Lake ; the Dead Sea,
·
Industrial plants generating salt by solar
evaporation of sea-water, and
·
Salted proteinaceous substances e.g., salted fish.**
In usual
practice, the colonies are found to
range from red to orange colouration by virtue of the presence of carotenoids*** that particularly appear
to cause adequate protection to the ensuing cells against the damaging effect
of the sunlight (having UV radiation).
Salient features : The
salient features of the Halobacterium and
the Halococcus cells are as stated below :
(1) The
cells do resist ‘dehydration’
particularly at high sodium chloride (NaCl) concentration due to the adequate
maintenance of a high intracellular osmotic concentration of potassium chloride
(KCl).
(2) Both
ribosomes and the cytoplasmic membrane are found to be fairly stable only at
relatively high concentrations of KCl, whereas the corresponding enzymes are
observed to be active only at high concentrations of either NaCl or KCl.
(3) Importantly,
the Halobacterium cell walls are
invariably made up of ‘certain protein
subunits’ which are held together only in the presence of NaCl ; and,
therefore, if the critical level of NaCl happens to fall below approximately
10% (w/v), the cells undergo break up.
(4) Interestingly,
the Halococcus cell walls are
usually comprised of a complex heteropolysaccharide which is found to
be stable reasonably at comparatively lower NaCl concentrations.
Adenosine Triphosphate (ATP) Synthesis. It is
worthwhile to mention here that generally the ‘halobacteria’ are ‘aerobic’
in nature. It is amply established that in
aerobic organisms, an
electron-transport chain invariably gives rise to a specific protonmotive force that in turn helps to carry out the desired ATP-Synthesis.
Salient Features : There are
several salient features that are associated with the ATP-synthesis, namely :
(1) ATP-synthesis
may alternatively be accomplished by halobacteria
via fermentation of arginine (an amino acid), which permits
them to grow in an anaerobic environment.
(2) The third method of ATP formation is rather
unique and extraordinary to the ‘halobacteria’.
Predominently distinct patches of a purple pigment, known as bacteriorhodopsin*, are pro-duced in
the cell membrane particularly at reasonably low O2 levels.
Subsequently, when these cells containing the said pigments are exposed to the
UV-light—the pigment gets bleached gradually. In the course of the ‘bleaching phenomenon’, the resulting
protons** get duly extruded right into the outside portion of the membrane,
thereby exerting an appre-ciable protonmotive force that in turn carries out
the ATP synthesis strategically.
(3) Conclusively,
halobacteria essentially follows the
mechanism of light-monitored synthesis of ATP. Furthermore, these are actually
devoid of bacteriochlorophyll.
3. Thermoacidophiles
The thermoacidophiles are generally the aerobic Gram –ve archaeobacteria
prominently char-acterized by a remarkable tendency and capability to attain
growth not only under extremely high
acidic conditions, but also at considerably elevated temperatures.
There are
two most prominent genera that belong
to this particular category, namely :
(a) Thermoplasma,
and
(b) Sulfolobus.
3.1 Thermoplasma
These
chemoorganotrophic microorganisms very much look alike the mycoplasm (i.e., a group
of organisms that lack cell walls and are highly pleomorphic), and obviously
varying from spherical in shape to filamentous in nature. The ideal and optimum
temperature for their progressive growth ranges between 55 and 59 °C (minimum,
44 °C ; maximum, 62 °C), whereas the optimum pH is 2 (minimum, 1 ; maximum, 4).
It has been duly observed that the cells of these thermoplasmas undergo abundant lysis virtually at a neutral pH. In
actual practice, the thermoplasmas
have been duly isolated from the re-sidual heaps of burning coal refuse.
3.2 Sulfolobus
The cells
of this particular genus are more or less lobe-shaped or spherical in shape and
appear-ance. They have the definite cell walls that are essentially made up of
protein. However, the optimum temperature and optimum pH of different species of sulfolobus are as given below :
Optimum
temperature : 70–87 °C ;
Optimum
pH : 2 [Min. 1 ; Max. 4].
Nevertheless,
the sulfolobus are established to be
autotrophic* facultatively. In fact,
sulfolobus may be grown in two different manners as stated under :
Method ‘A’ — as ‘chemolithotrophs’ when adequately provided with ‘S’ as
an element and an electron donor,
and
Method ‘B’ — as ‘chemoorganotrophs’ in the respective media comprising of
organic substrates.
Interestingly,
the natural occurrence of the sulfolobus
species are prominently and predominently found in sulphur (acidic) hot springs
around the world.
Related Topics