Types of Microbial Cells
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Structure and Function of Bacterial Cells
As to date there are two types of cells that have been recognized duly, such as : (a) Eukaryotic cells, and (b) Prokaryotic cells.
Types of Cells
As to date there are two types of cells that have been recognized duly, such as :
(a) Eukaryotic cells, and
(b) Prokaryotic cells.
Another third type, known as the Urkaryotes, and are most probably the progenitor of the present day eukaryotes has now also been recognized duly.
The above two types of cells* (a) and (b) shall now be discussed at length in the sections that follows :
(a) Eukaryotic Cells [‘eu’ = true ; ‘karyote’ = nut (refers to nucleus of cell)] ;
It has been observed that the eukaryotic cells (Fig. 2.1) are explicitely characterized by the presence of a multiplicity of definite unit membrane systems that happen to be both structurally and topologically distinct from the cytoplasmic membrane. Subsequently, these prevailing membrane sys-tems categorically enable the segregation of various eukaryotic cytoplasmic functions directly into specialized organelles.** Endoplasmic reticulum (ER) represents the most complex internal mem-brane system that essentially comprises of an irregular network of interconnected delimited channels that invariably cover a larger segment of the interior portion of the cell. Besides, ER gets in direct contact with two other extremely vital components viz., nucleus and cytoplasmic ribosomes. The nu-cleus membrane is duly formed by a portion of the endoplasmic reticulum surrounding the nucleus ; whereas, in other regions the surface of the membrane is particularly covered with the ribosomes wher-ever synthesis of protein takes place. The proteins thus generated eventually pass via the endoplasmic reticulum channels right to the various segments of the ensuing cell cytoplasm.
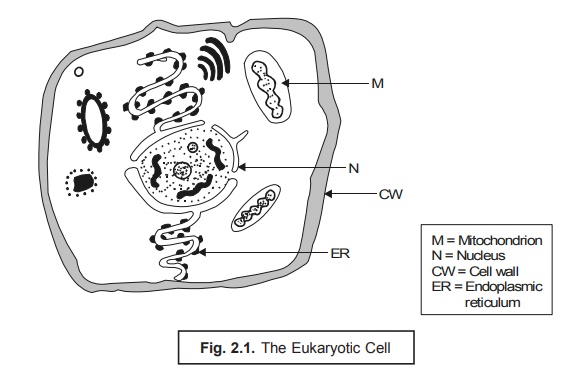
Nucleus. The eukaryotic cell possesses the ‘genetic material’ duly stored in the chromosomes i.e., very much within the nucleus. However, chloroplasts and mitochondria also comprise of character-istic DNA. The chromosomes are linear threads made of DNA (and proteins in eukaryotic cells) in the nucleus of a cell, which may stain deeply with basic dyes, and are found to be especially conspicuous during mitosis. The DNA happens to be the genetic code of the cell ; and specific sequences of DNA nucleotides are the genes for the cell’s particular proteins. However, the size and the number of the chromosome vary widely with various organisms. Nevertheless, the nucleus invariably contains a nu-cleolus that is intimately associated with a particular chromosomal segment termed as the ‘nucleolar organizer’, which is considered to be totally involved in ribosomal RNA (rRNA) synthesis.
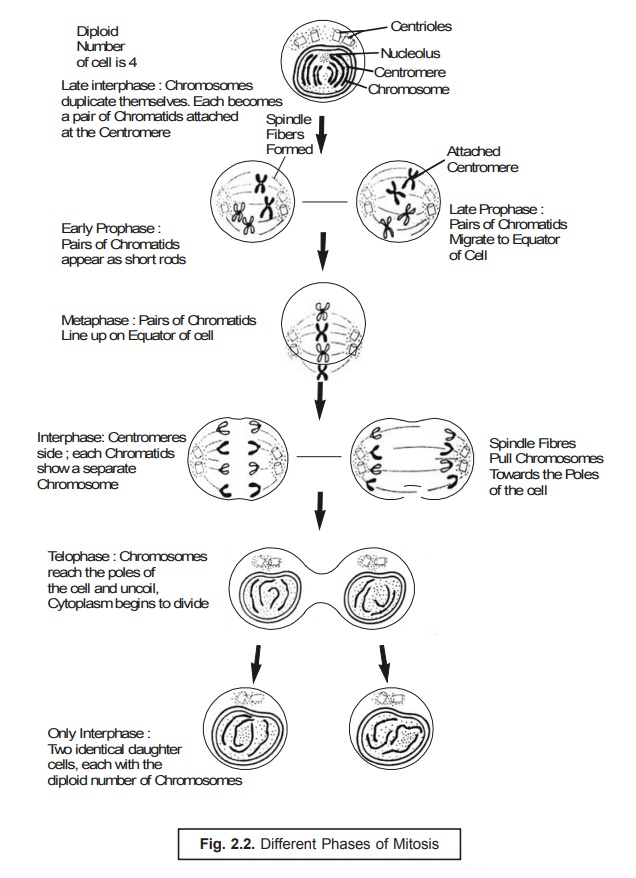
Mitosis. Mitosis refers to a type of cell division of somatic cells wherein each daughter cell contains the same number of chromosomes as the parent cell. Mitosis is the specific process by which the body grows and dead somatic cells are replaced. In fact, mitosis is a continuous process divided into four distinct phases, namely : prophase, metaphase, anaphase, and telophase.
A brief discussion of the aforesaid four phases shall be given in the sections that follows along with their illustrations in Fig. 2.2.
(a) Prophase. In prophase, the chromatin granules of the nucleus usually stain more densely and get organized into chromosomes. These first appear as long filaments, each comprising of two identical chromatids,* obtained as a result of DNA replication. As prophase progresses, the chromosomes become shorter and more compact and stain densely. The nuclear mem-brane and the nucleoli disappear. At the same time, the centriole divides and the two daugh-ter centrioles,** each surrounded by a centrosphere, move to opposite poles of the cell. They are duly connected by fine protoplasmic fibrils, which eventually form an achromatic spindle.
(b) Metaphase. The metaphase refers to the chromosomes (paired chromatids) that arrange themselves in an equatorial plane midway between the two centrioles.
(c) Anaphase. In anaphase, the chromatids (now known as daughter chromosomes) diverge and move towards their respective centrosomes. The end of their migration marks the begin-ning of the next phase.
(d) Telophase. In telophase, the chromosomes at each pole of the spindle undergo changes that are the reverse of those in the prophase, each becoming a long loosely spiraled thread. The nuclear membrane re-forms and nucleoli reappear. Outlines of chromosomes disappear, and chromatin appears as granules scattered throughout the nucleus and connected by a highly staining net. The cytoplasm gets separated into two portions, ultimately resulting in two complete cells. This is accomplished in animal cells by constriction in the equatorial region ; in plant cells, a cell plate that produces the cell membrane forms in a similar position. The period between two successive divisions is usually known as interphase.
Mitosis is of particular significance wherein the genes are distributed equally to each daughter cell and a fixed number of chromosomes is maintained in all somatic cells of an organism.
Mitosis are of two kinds, namely :
(i) heterotypic mitosis : The first or reduction division in the maturation of germ cells, and
(ii) homeotypic mitosis : The second or equational division in the maturation of germ cells.
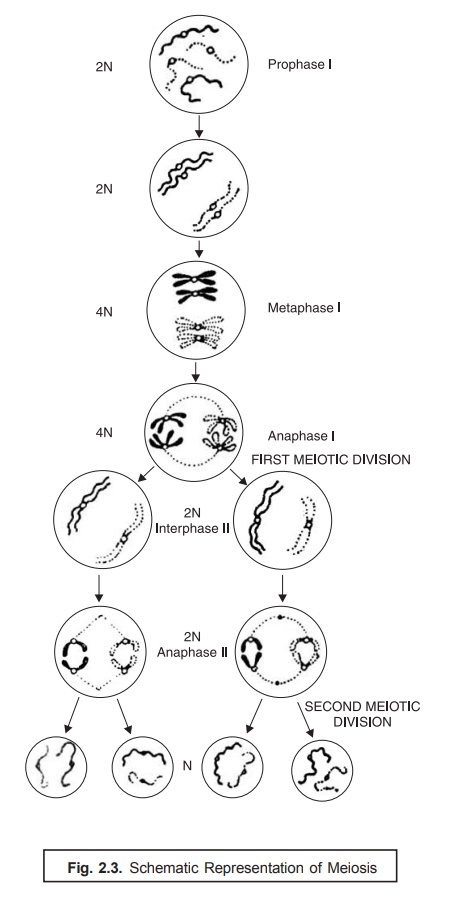
Meiosis. Meiosis refers to a specific process of two successive cell divisions, giving rise to cells, egg or sperm, that essentially contain half the number of chromosomes in somatic cells. When fertiliza-tion takes place, the nuclei of the sperm and ovum fuse and produce a zygote with the full chromosome complement.
In other words, the phenomenon of meiosis may be duly expatiated in sexually reproducing organisms, wherein the prevailing cellular fusion followed by a reduction in the ‘chromosome number’ is an important and vital feature. The two cells which actually participate in the sexual reproduction are termed as ‘gametes’, which fuse to form a ‘zygote’. The above process is subsequently followed by a nuclear fusion and the resulting zygote nucleus contains two complete sets of genetic determinants [2N]. In order to adequately maintain the original haploid number in the succeeding generations, there should be a particular stage at which a definite reduction in the chromosome number takes place. This process that occurs after the fusion of gametes is known as meiosis.
Fig. 2.3 illustrates the schematic representation of meiosis, and the various steps involved may be explained sequentially as follows :
(1) Meiosis comprises of two meiotic divisions viz., prophase I, and prophase II.
(2) Prophase-I. It represents the first meiotic division, whereby the homologous chromosomes become apparently visible as single strands that subsequently undergo pairing.
(3) Each chromosome renders visible as two distinct chromatids and thus crossing over takes place.
(4) It is immediately followed by metaphase I, wherein the actual orientation of ‘paired chro-mosomes’ in an equatorial plane and the subsequent formation of a ‘spindle apparatus’ takes place.
(5) It is followed by Anaphase I, and the homologous centromeres gradually move to the opposite poles of the spindle.
(6) Telophase-I. It markedly represents the end of the first meiotic division, and formation of two nuclei takes place.
(8) Interphase-II. Telophase-I is followed by Interphase-I during which the chromosomes get elongated.
(8) Prophase-II and Metaphase-II. In prophase-II and metaphase-II the division of centromere and migration of the homologous chromatids occurs, which is duly followed by anaphase-II, and the desired second meiotic division resulting in the formation of four haploid* cells.
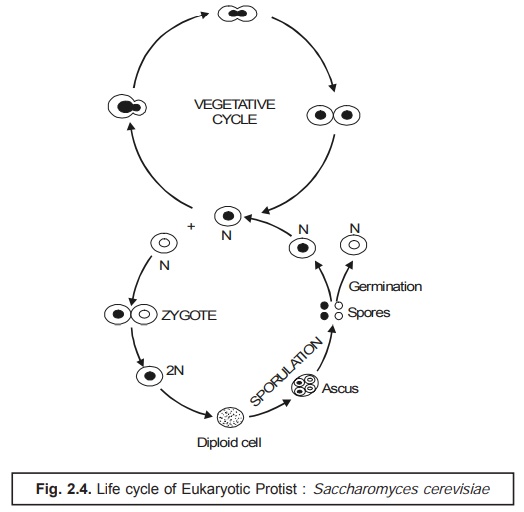
Eukaryotic Protist. It has been observed that in several eukaryotic protists belonging to higher ploidy** (> 1) meiosis usually takes place after the formation of the zygote and prior to spore formation. In certain eukaryotes there may even be a critically pronounced alteration of haploid and diploid gen-erations as in the case of the yeast. Interestingly, in this particular instance, the diploid zygote produces a diploid individual that ultimately gives rise to haploid cells only after having undergone the phenom-enon of meiosis. Consequently, the haploid cell may either multiply as a haploid or get fused with another haploid of the ‘opposite mating type’ to generate again a diploid.
Example. The life cycle of the eukaryotic protist may be exemplified by a typical yeast Saccharomyces cerevisae as depicted in Fig. 2.4 given below :
Special Points : There are two cardinal points which, may be borne in mind with regard to the Eukaryotic Protist as stated under :
(i) Despite of the fact that sexual reproduction could be the only way of reproduction in a large segment of animals and plants ; it may not be an obligatory event in the life cycles of many protists.
(ii) In two glaring situations ; first, protists lacking a sexual stage in their respective life-cycle ; and secondly, such species wherein sexuality does exist : the sexual reproduction may be quite infrequent (i.e., not-so-common).
Important organelles in Eukaryotic Cells : It has been amply proved and established that the eukaryotic cells invariably contain certain cytoplasmic organelles other than the nucleus. The important organelles in eukaryotic cells usually comprise of three components, namely : mitochondria, chloroplasts, and the Golgi apparatus, which shall now be described briefly in the sections that follows :
Mitochondria. These are mostly found in the respiring eukaryotes and essentially contain an internal membrane system having characteristic structure and function. The internal membrane of the mitochondria (cristae) possesses the necessary respiratory electron transport system. The exact number of copies of mitochondria per cell solely depends upon the cultural parameters and varies from 1–20 mitochondria per cell. These are generated by the division of the preexisting organelles containing ribosomes that usually resemble the bacterial ribosomes. However, the process of protein synthesis in the mitochondria are very much akin to that in the prokaryotic cells.
These cell organelles (rod/oval shape 0.5 μm in diameter) may be seen by employing a phase-contrast or electron microscopy. They mostly contain the enzymes for the aerobic stages of cell respi-ration and thus are the usual sites of most ATP synthesis chloroplasts [or Chloroplastids] :
Chloroplasts are found in the photosynthetic eukaryotic organisms. The internal membrane of the chloroplasts is termed as the ‘thylakoid’ which essentially has the three important components : (a) photosynthetic pigments, (b) electron transport system, and (c) photochemical reaction centres. The number of copies of the chloroplasts depends exclusively upon the cultural conditions and varies from 40 to 50 chloroplasts per cell. These are also produced by the division of the preexisting organelles.
Generally, chloroplasts are the sites of photosynthesis. They possess a stroma and contain four pigments : chlorophyll a, chlorophyll b, carotene, and xanthophyll.
Golgi Apparatus : The Golgi apparatus is a lamellar membranous organelle invariably found in the eukaryotic cells and consists of thickly packed mass of flattened vessels and sacks of different sizes. The major functions of the Golgi apparatus are, namely :
· packaging of both proteinaceous and nonproteinaceous substances duly synthesized in the endoplasmic reticulum, and
· their adequate transport to other segments of the cell.
Golgi apparatus may be best viewed by the aid of electron microscopy. It contains curved parallel series of flattened saccules that are often expanded at their ends. In secretory cells, the apparatus concentrates and packages the secretory product. Its function in other cells, although apparently impor-tant, is poorly understood.
(b) Prokaryotic Cells [‘pro’ = primitive ; ‘karyote’ = nut (refers to nucleus of cell)] :
Prokaryote : is an organism of the kingdom Monera with a single circular chromosome, without a nuclear membrane, or membrane bound organelles. Included in this classification are bacteria and cyanobacteria (formerly the blue-green algae) [SYN : prokaryote].
In fact, the prokaryotic cell is characterized by the absence of the endoplasmic reticulum (ER) and the cytoplasmic membrane happens to be the only unit membrane of the cell. If has been observed that the cytoplasmic membrane may be occasionally unfolded deep into the cytoplasm. An exhaustive electron microscopical studies would reveal that most prokaryotes {i.e., prokaryotic cells) only two distinct internal regions, namely : (a) the cytoplasm ; and (b) the nucleoplasm, as shown in Fig. : 2.5.
Cytoplasm : Cytoplasm refers to the protoplasm cell outside the nucleus. It is granular in ap-pearance and contains ribosomes that are specifically smaller in size in comparison to the corresponding eukaryotic ribosomes.
Nucleoplasm : It refers to the protoplasm of a cell nucleus. It is fibrillar in character and contains DNA.
With mycoplasmas* as an exception, other prokaryotes invariably comprise of a defined and rigid cell wall. It has been observed that neither the membranous structures very much identical to the mitochondria nor chloroplasts are present in the prokaryotes. Besides, the cytoplasmic membrane happens to be the site of the respiratory electron in the prokaryotes usually. Interestingly, in the photosynthetic micro- organisms (bacteria), the photosynthetic apparatus is strategically positioned in a particular series of membranous, flattened structures quite similar in appearance to the thylakoids ; however, these struc-tures are not organized into the respective chloroplasts but are adequately dispersed in the cytoplasm. Thus, the cytoplasmic membrane contains a plethora of specific sites for the DNA attachment, and also plays a major role in the cell division. Here, the cell membrane unlike in the eukaryotic cell does not generally contain sterols and polyunsaturated fatty acids (PUFAs). Mostly the fatty acids present are of the saturated type e.g., palmitic acid, stearic acid etc.
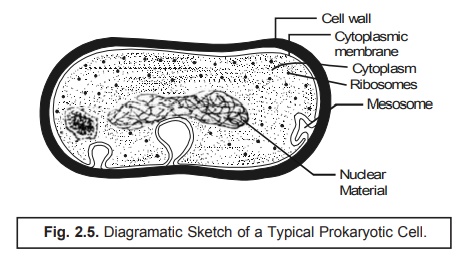
Importantly, the ‘genetic component’ present in the prokaryotic cells is strategically located in the ‘nucleoplasm’ that essentially lacks a defined nuclear membrane. Nevertheless, it comprises of dou-ble helical DNA without any associated basic proteins. In fact, the very site of the DNA in prokaryotic protists is much smaller in comparison to that present in eukaryotes. In addition, the prokaryotes do contain extra-chromosomal DNA, that may replicate autonomously, termed as the ‘plasmids’. How-ever, these can be lost from the cell without impairment of the ‘cell viability’. The prokaryotic cells usually exist in a haploid state and predominently get divided by a process quite identical to mitosis although distinct stages are not recognized so frequently.
A good number of prokaryotes do possess a cell wall that is vastly different in composition from that of eukaryotes, and invariably contains a rather rigid and well-defined polymer termed as the peptidoglycan.* It has been observed that certain prokaryotes which essentially possess this aforesaid rigid structure distinctly exhibit ‘active movement’ with the help of flagella. Some prokaryotes may also display a ‘gliding motility’ as could be seen in the ‘blue-green bacteria’ quite frequently.
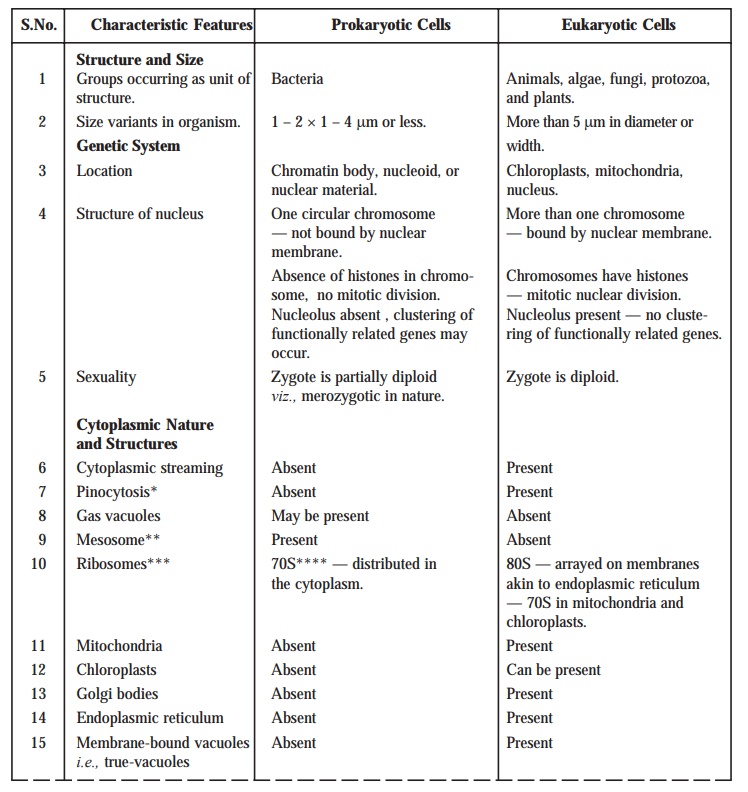
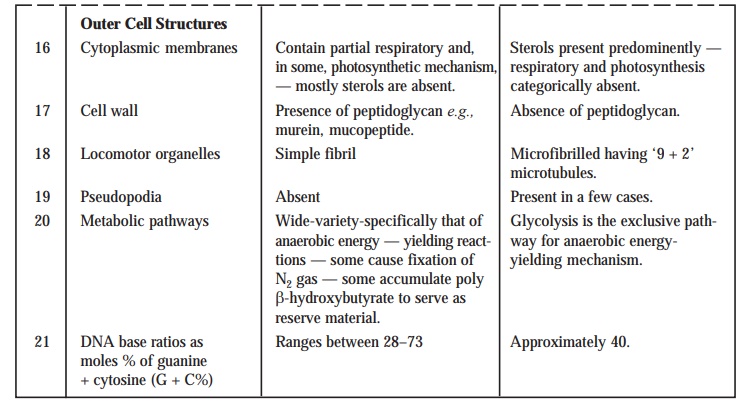
Table : 2.2. records the distinguishing characteristic features of the Prokaryotic from the Eukaryotic Cells.
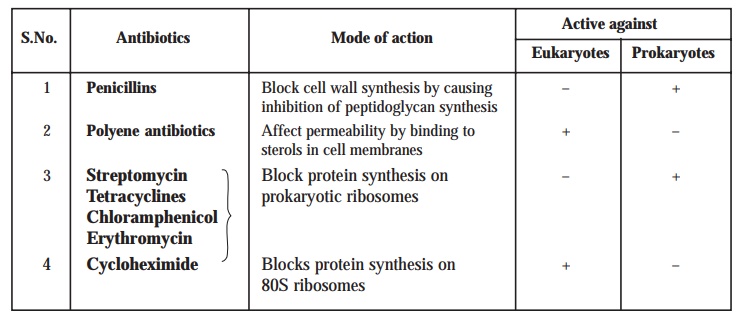
Selective sensitivity to antibiotics. Another reliable and practical means to differentiate the eukaryotes from prokaryotes is their characteristic selective sensitivity to certain specific antibiotic(s). However, one may observe that chloramphenicol is toxic only to bacteria, whereas polyene antibiotics (e.g., nystatin) bind to sterols in the cell membranes, and are largely effective exclusively against the eukaryotic protists.
Table 2.3 : summarizes actually the vital and important differences in the activity against the eukaryotes and prokaryotes with respect to selective sensitivity to ‘antibiotics’ vis-a-vis their mode of action.
Table 2.3. Differences Between Eukaryotes and Prokaryotes as Regards Selective Sensitivity to Antibiotics/Mode of Action
It is, however, pertinent to mention here that several cellular functionalities are prominently and predominently mediated almost differently in these two distinct types of cells, although the end result is more or less the same.
Related Topics
