Structure and Function of Bacterial Cells
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Structure and Function of Bacterial Cells
The present section shall encompass briefly the major cellular structures usually encountered in the bacteria. Nevertheless, the various functional anatomy of these cell types would throw an ample light upon the various special activities that such cells perform normally.
THE
BACTERIAL CELLS
The
present section shall encompass briefly the major cellular structures usually
encountered in the bacteria. Nevertheless, the various functional anatomy of
these cell types would throw an ample light upon the various special activities
that such cells perform normally.
The
cellular structure should essentially provide the following three cardinal objectives, namely :
(a) a
specific container to support the internal contents and to segregate it totally
from the exter-nal medium,
(b) to
store and replicate the genetic information, and
(c) to
synthesize energy and other necessary cellular components for the replication
of the cell.
In
general, the bacterial cells grossly fulfil these requirements completely ;
besides, they have distinguishable characteristic features to help
differentiation one from the other.
It is,
however, pertinent to state here that extensive hurdles and difficulties were
encountered by the microbiologists across the globe in carrying out the
detailed cytological studies** of
bacteria on account of the following vital factors, such as :
(i) the extremely small size
(dimension) of the microorganism, and
(ii) almost optically homogeneous nature
of the cytoplasm.
As to
date, the advent of the development of complex and precisely selective staining
techniques amalgamated with the magnificent discovery of electron microscope and phase-contrast
microscope have contributed enormously in obtaining a far better in-depth
knowledge and understanding of the ‘internal
structures of bacterial cells’.
The
various important aspects referring to the domain of the ‘bacterial cells’ shall be adequately dealt with under the
following heads stated as under :
(i) Typical
bacterial cell
(ii) Capsules
and slimes
(iii) Flagella
and fimbria
(iv) Cell
envelope
(v) Gram-positive
and gram-negative bacteria
(vi) Significance
of teichoic acids
(vii) The
cell membrane
(viii) Bacterial
cytoplasm
(ix) Ribosomes,
and
(x) Cellular
reserve materials
1. Typical Bacterial Cell
Bacteria being prokaryotic in nature are much simpler in
comparison to the ‘animal cells’. In addition
to this, they have three distinct
characteristic features, namely : (a)
an extensive endoplasmic reticulum* ; (b)
essentially lack a membrane-bound nucleus ; and (c) mitochondria.
Nevertheless,
bacteria do possess a rather complex
surface structure having a rigid cell
wall that surrounds the cytoplasmic
membrane, as shown in Fig. 2.7, which essentially serves as the osmotic barrier as well as the ‘active transport’ necessarily needed so as
to sustain and maintain a suitable intracellular
concentration of the specific ions and the metabolites.
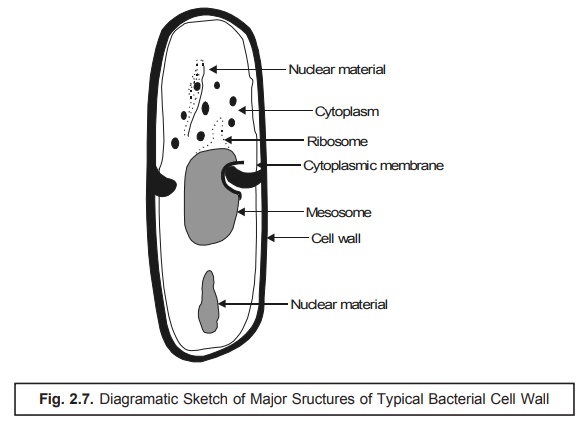
Infact,
the bacterial cell wall has two major roles to play :
(a) to
protect the cell against osmotic rupture particularly in diluted media, and
also against certain possible mechanical damage(s), and
(b) to
assign bacterial shapes, their subsequent major division into Gram positive and
Gram negative microorganisms and their antigenic attributes.
2. Capsules and Slimes
Invariably
certain bacterial cells are duly
surrounded by a viscous material that essentially forms a covering layer or a
sort of envelope around the cell wall. In the event this specific layer may be visualized by the aid of light microscopy
employing highly sophisticated and specialized staining tech-niques, it is
known as a capsule; in case, the layer happens to be too thin to be
observed by light microscopy, it is called as a microcapsule. If the layer
does exist in an absolute abundance such that quite many cells are found to be
embedded in a common matrix, the
substance is termed as a slime.
In other
words, the terminology capsule
usually refers to the layer both intimately and tightly attached to the cell
wall ; whereas, the slime coating
(layer) is contrarily the loose structure which often gets diffused right into
the corresponding available growth medium as depicted in Fig. 2.8 below :
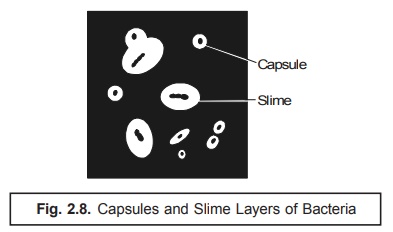
Salient features : The
salient features of capsule and slime are
enumerated as under :
(1) These
structures are not quite necessary and important for the normal growth and
usual survival of the bacterial cells
but their very presence grants some apparent advantages to the bacterial cells that contain these
structures.
(2) A
plethora of bacteria are incapable of producing either a capsule or a slime ; and
those which can do so would certainly lose the ability to synthesize
legitimately these two compo-nents devoid of any adverse effects.
(3) The
prime interest in these amorphous organic
exopolymers i.e., capsules and slimes, was to assess precisely their actual
role in the pathogenicity by virtue of the fact that majority of these
pathogenic microorganisms do produce either a capsule or a slime.
It is
worthwhile to mention here that the composition of such amorphous organic exopolymers varies according to the particular
species of bacteria. In certain
instances these are found as homopolymers
essentially of either carbohydrates (sugars) or amino acids, whereas in
other cases these could be seen as heteropolymers essentially of
carbohydrates/substituted carbohydrates e.g.,
heteropolysaccharides.
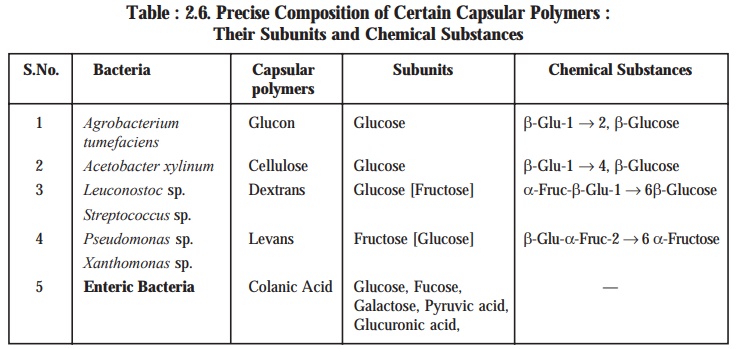
A few
typical examples of specific microorganisms
(bacteria) having a varied range of amor-phous organic exopolymers are as
given below :

Further
investigative studies on different types of organisms (bacteria) have revealed,
the precise composition of a few selective capsular polymers (i.e., amorphous organic exopolymers)
along with their respective subunits and chemical substances produced at the
end, as provided in Table 2.6.
Table : 2.6. Precise Composition of Certain
Capsular Polymers : Their Subunits and Chemical Substances
Important Points : There
are five important points that may be noted carefully :
(i) It is
still a mystery to know that on one hand in certain bacteria the exopolymers
are seen in the form of capsules ; whereas, on the other they are observed in
the form of slimes.
(ii) Mutation*
of capsular form to the corresponding slime forming bacteria has been well
established.
(iii) Structural
integrity of both the capsule as well as the slime are meticulously estimated
by the critical presence of distinct chemical entities.
(iv) In
many cases, the capsular material is not extremely water-soluble ; and,
therefore, fails to diffuse rapidly away from the cells that eventually produce
it.
(v) In
certain other instances the capsular material is highly water-soluble ; and
hence, either gets dissolved in the medium instantly or sometimes abruptly
enhancing the viscosity of the broth in which organisms are cultured
respectively.
Functions of Capsules : In
reality capsules may serve five cardinal functions exclusively
de-pending upon their respective bacterial species as described under:
(a) They
may afford adequate protection against temporary drying by strategically bound
to water molecules.
(b) They
may cause absolute blockade of attachment to bacteriophages.
(c) They
may be antiphagocytic* in nature.
(d) They
may invariably promote attachment of bacteria to surfaces, such as : Streptococcus mutans — a bacterium that is directly linked to causing dental
caries, by means of its ability to adhere intimately onto the smooth
surfaces of teeth on account of its specific secretion of a water-insoluble capsular glucan.
(e) In
the event when the capsules are essentially made up of compounds bearing an ‘electrical charge’, for instance: a combination of sugar-uronic acids, they may duly help in the pro-motion of
the stability of bacterial suspension
by preventing the cells from aggregating and settling out by virtue of the fact
that such cells having identical charged surfaces would have a tendency to
repel one another predominently.
3. Flagella and Fimbria
3.1. Flagella
Flagellum [Pl : Flagella] refers to a thread like structure that provides
motility for certain bacte-ria and protozoa (one, few or many per cell) and for
spermatazoa (one per cell).
It has
been observed that the presence of flagella
strategically located on certain bacteria
(miroorganisms) has been known ever since the beginning of the nineteenth
century ; besides, the actual form of flagellation and motility have been
exploited judiciously as a taxonomic tool in the logical classification of
bacterial variants.
Filaments : The ‘flagella’ are nothing but surface appendages
invariably found in motile bacte-ria, and
appear generally as filaments having
diameter ranging between 12–20 nm and length between 6–8 μm.
Importantly, the diameter of the individual flagellum in a culture is normally
constant ; how-ever, the length may vary accordingly.
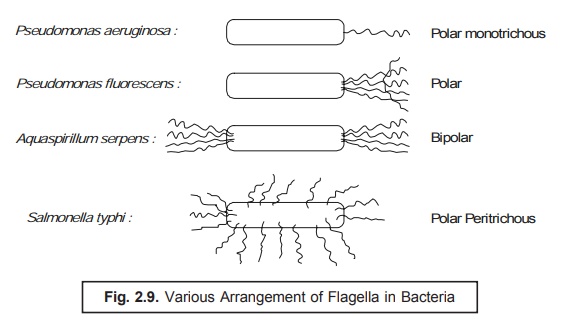
Location of Flagella : The exact location of the flagella in various
bacteria varies widely and specifically
; and could be either polar monotrichous
or polar or bipolar or polar
peritrichous as shown in Fig. 2.9 ; and the number of flagella per cell
also changes with the various bacterial species.
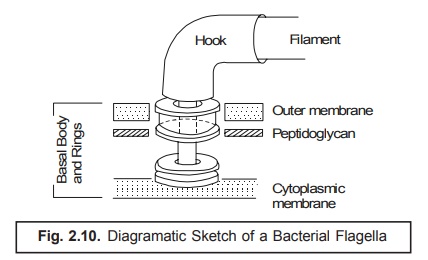
Flagellar Apparatus : Basically
the flagellar apparatus consists of three distinct parts, namely : (a)
filament ; (b) hook ; and (c) basal granule. Importantly, the
outermost structural segment of bacteria is the filament which is a fibre essentially comprised of a specific
protein termed as flagellin (a
subunit having molecular weight 20,000), and this is securedly attached to the basal granule with the help of the hook.
Interestingly,
both the basal granules and the hook essentially contain certain
specific proteins that are antigenically
distinct from the flagellin (i.e., the protein of the filament).
In fact,
the particular structure of the basal
body comprises of a small central rod inserted strate-gically into a system
of rings as illustrated in Fig. 2.10 below. However, the entire unit just functions
fundamentally as a ‘simple motor’. It
has been amply demonstrated and established that the meticulous growth of the
flagella invariably takes place by the careful addition of the flagellin subunits at the distal end
after being drifted through from the cytoplasm, obviously via the hollow core of the very flagellum.
Functioning of Flagella : The modus
operandi of flagella are as given under :
(1) Flagella
are fully responsible for the bacterial motility.
(2) Deflagellation
by mechanical means renders the motile cells immotile.
(3) The
apparent movement of the bacterial cell usually takes place by the critical
rotation of the flagella either in the clockwise or anticlockwise direction
along its long axis.
(4) Bacterial
cell possesses the inherent capacity to alter both the direction of rotation
[as in (3) above] and the speed ; besides, the meticulous adjustment of
frequency of ‘stops’ and ‘starts’ by the appropriate movement of the flagella.
(5) Evidently,
the flagellated peritrichal* bacteria usually swim in a straight line over
moderate distances. In actual practice, these swim-across straight line runs
are interrupted frequently by abrupt alterations in the direction that
ultimately leads to tumbling. Therefore, the move-ment of the bacteria is
believed to be zig-zag.
(6) It
has been observed that the phenomenon of smooth swimming in a fixed direction
is invari-ably mediated by the rotation of flagella in an anticlockwise
direction ; whereas, the process of tumbling in a zig-zag direction is usually
caused by the rotation of flagella in a clockwise direction.
(7) The
presence of ‘polar flagella’ in
bacteria affords a distinct change in the direction that usually takes place by
the reciprocal alteration in the direction of rotation.
3.2. Fimbriae [or Pili*]
Fimbriae or Pili are hollow, non-helical, filamentous hair-like structures
that are apparently thinner,
shorter, and more numerous than flagella. However, these structures do appear
on the surface of the only Gram negative bacteria and are virtually distinct
from the flagella.
Another
school of thought rightly differentiates the terminology ‘fimbriae’ exclusively reserved for all hair-like structures ;
whereas, other structures that are directly and intimately involved in the
actual transfer of genetic material solely are termed as ‘pili’. Likewise, the bacterial
flagella that may be visualized conveniently with the help of a light microscope after only suitable
staining ; and the bacterial pili can
be seen vividly only with the aid of an electron microscope.
Salient Features of Fimbriae : Some of
the important salient features of
‘fimbriae’ are as enumerated
under :
(1) At
least 5 to 6 fimbriae variants have been recognized besides the sex pili.
(2) Type I fimbriae has been
characterized completely.
(3) They
contain a particular protein known as pilin
having molecular weight of 17,000 daltons.
(4) The
fimbriae are found to be spread over the entire cell surface. These have a
diameter of 7 nm and a length ranging between 0.5 to 2 μm ; besides, an empty core of 2
to 2.5 nm.
(5) The pilin subunits are duly arranged in a
helical manner having the pitch of the helix** almost nearly at 2.3 μm.
(6) In
addition to the Type-I fimbriae, the
Gram-negative bacteria invariably own a special category of pili termed as the sex pili (or F-pili), the synthesis of which is predominently directed by the sex factor (or F-factor). It has been observed that the sex pili do have a uniform diameter of approximately 9 nm, and a
length almost nearing between 1-20 μm.
(7) Very
much akin to the flagella, both fimbriae and pili are observed to originate from the basal bodies strategically located within the cytoplasm. Interestingly, neither fimbriae nor
pili seem to be essential for
the survival of the bacteria.
Human Infection : It has
been demonstrated that certain pili do
play a major role in causing and spreading
human infection to an appreciable
extent by permitting the pathogenic bacteria to get strategically attached to
various epithelial cells lining the genito urinary, intestinal, or respiratory
tracts specifically. It is worthwhile to mention here that this particular
attachment exclusively checks and pre-vents the bacteria from being washed away
critically by the incessent flow of either mucous or body fluids thereby
allowing the infection to be established rather firmly.
4. Cell Envelope
Extensive
morphological investigations have adequately revealed that the cell envelope of the Gram-positive bacteria* is much more
simpler with regard to the structure in comparison to that of the Gram-negative bacteria.**
For Gram-positive Bacteria : In this
instance the cell envelope contains chiefly the peptidoglycan and the teichoic acids.
Interestingly,
the peptidoglycan represents a substituted carbohydrate polymer found
exclusively in the prokaryotic
microorganisms.
It
essentially comprises of two major
chemical entities namely :
(a) Two acetylated aminosugars e.g., n-acetyl
glucosamine ; and n-acetylmuramic
acid ; and
(b) Amino acids e.g.,
D-glutamic acid ; D- and L-alanine ;
In fact,
the long peptide chains containing
the two amino sugars that essentially constitute the ‘glycan strands’ comprise of alternating units of n-acetyl
glucosamine and n-acetyl muramic acid in β-1, 4-linkage ; besides, each strand
predominently contains disaccharide residues ranging from 10 to 65 units
as shown in Fig. 2.11.
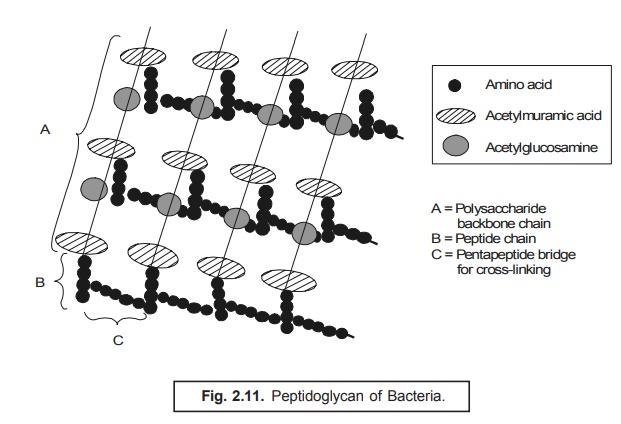
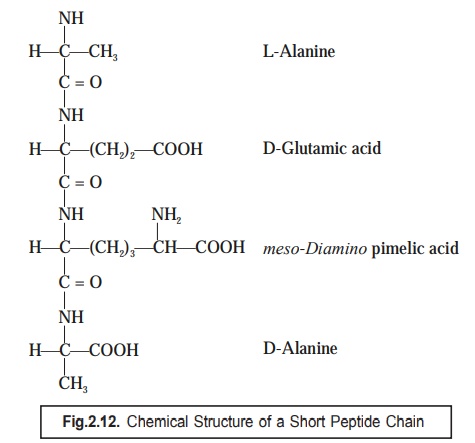
Nevertheless,
the short peptide chains consisting
four amino acids are found to be strategically linked to the corresponding muramic acid residues ; and invariably
the most commonly encountered sequence being L-alanine, D-glutamic acid, meso-diamino pimelic acid, and
D-alanine, as depicted in Fig. 2.12.
Salient Features : The
various important and noteworthy salient features with regard to the formation of peptide chains are as
enumerated under :
(1) The
3rd amino acid i.e., meso-diamino
pimelic acid (Fig. 2.12) has been observed to vary with different organisms (bacteria)
by any one of the three such amino
acids as : lysine, diamino pimelic acid, or threonine.
(2) Besides,
the adjacent peptide chains occurring in a peptidoglycan
could be duly cross-linked by short peptide chains essentially comprising of a
varying number of amino acids.
(3) An
important characteristic feature viz.,
the variations in the structure of the peptidoglycan
constituents usually take place ; and, therefore, it has been exploited and
utilized judiciously as a wonderful
taxonomic tool.
(4) The
exact number of amino acids that eventually form the cross link prevailing
between the two n-acetyl muramic acid
residue i.e., the interpeptide bridge
variation, may also vary from 2 to 5,
as given in Table 2.7, depending upon the various species of microorganisms.
Varia-tions in the n-acetyl muramic acid are also known and these alterations
ultimately do affect the compactness of the peptidoglycan
to an appreciable degree.

5. Gram-Positive and Gram-Negative Bacteria
The various
characteristic features of Gram-positive and Gram-negative bacteria shall be
dis-cussed at length in this particular section.
For Gram-negative bacteria. There are two distinct layers that have been duly
recognized in the cell envelopes of Gram-negative bacteria, namely :
(a) An
uniform inner layer approximately 2–3
mm wide, and
(b) A
thicker outer layer nearly 8–10 nm
wide.
Importantly,
the peptidoglycan is prominently
confined to the inner layer ; whereas, the outer layer (membrane) essentially comprises
of proteins, lipoproteins, and lipopolysaccharides.
The
principal chemical differences that predominently occur between the cell walls
of Gram-positive bacteria and the inner rigid wall layer and outer wall
layer(s) of Gram-negative bacteria have been duly summarized in Table 2.8 given
below :
Table 2.8. Principal Chemical Differences Existing
Between Cell Walls of Gram-positive and Gram-negative Bacteria

Cardinal characteristic features of component
variants in Gram +ve and Gram –ve micro-organisms : The
various important characteristic features of component variants in Gram +ve and
Gram –ve microbes are as stated
under :
(1) Peptidoglycans
belonging to the Gram –ve microorganisms exhibits a rather low extent of cross
linkages within the glycan strands.
(2) Outer-membrane. The fine
structure of the outer membrane, very much akin to cell mem-brane, essentially comprises of a lipid
bilayer wherein both phospholipids and lipopolysaccharides are definitely
present. Besides, the lipopolysaccharide generates the major component of the
outer membrane, and represents an extremely complex molecule varying in
chemical composition within/between the Gram –ve bacteria.
(3) Outer surface. The
peptidoglycan of the wall has particular kinds of lipoproteins residing on its outer surface, that are strategically linked by peptide bonds to
certain diaminopimelic acid residues present in the peptidoglycan.
(4) Lipoproteins
evidently serve as a sort of bridge right from the peptidoglycan upto the
outer-wall-layer.
(5) The
total number of proteins definitely present, unlike in the inner membrane, are
quite a few in number (approx. 10) ; and, therefore, these are markedly
distinct from those invari-ably found in the inner membrane.
Typical Example : It has
been observed that the lipopolysaccharides belonging to either E. coli or Salmonella sp.
necessarily comprise of subunits,
and each subunit consists of three
vital compo-nents, namely : (a) a
lipid ; (b) core region ; and (c) O-side chain respectively, as given in Fig.
2.13.
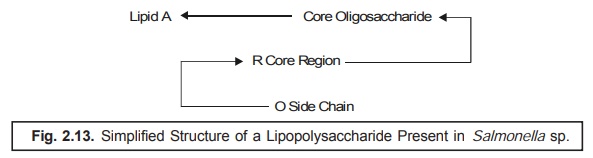
Explanations : The proper explanations for the
various transformations occurring in Figure : 2.13 are as given below :
(i) The
various subunits in lipopolysaccharide are duly linked via pyrophosphates with
the ‘lipid zone’.
(ii) The
‘lipid zone’ comprises of a phosphorylated glucosamine disaccharide esterified
adequately with long chain fatty acids.
(iii) The
‘core region’ comprises of a short-chain of carbohydrates, and the O-side chain
consists of different carbohydrates and is much longer in comparison to the
R-core region.
(iv) Lipopolysaccharides
represent the major antigenic determinants, and also the receptors for the
active adsorption of several bacteriophages.
Comparative Activities of Gram-negative and Gram-positive Bacteria
The
various glaring comparative activities of both Gram-negative and Gram-positive
bacteria are enumerated below :
(1) It
has been duly demonstrated that the outer membrane of Gram-negative bacteria
promi-nently behaves as a solid barrier to the smooth passage of certain
critical substances, for instance : antibiotics, bile salts*, and dyes into the
cell. Hence, the Gram-negative organisms are comparatively much less sensitive
to these substances than the Gram-positive ones.
(2) Adequate
treatment of Gram-negative bacteria with an appropriate chelating agent, such
as : ethylenediaminetetra acetic acid (EDTA), that eventually affords the
release of a substantial amount of lipopolysaccharides
renders ultimately the cells more sensitive to the drugs and chemical entities.
Thus, the presence of lipopolysaccharide
on the surface of the cell also helps the bacteria to become resistant to the phagocytes** of the host.
(3) The
resistance acquired in (2) above is almost lost only if the host enables to
synthesize the antibodies that are
particularly directed against the O-side chain (Figure 2.13). There exists a vast diversification in the specific
structure of the O-side chain ; and, therefore, gives rise to the somatic*** antigenic specificity very
much within the natural bacterial
populations. Evidently, the ensuing antigenic
diversity exhibits a distinct
selective advantage specifi-cally for a pathogenic bacterial species,
because the animal host is not in a position to pos-sess higher antibody levels
strategically directed against a relatively large number of varie-ties of
O-side chains.
(4) In
general, the prevailing lipids are invariably found to be phosphatidylethanolamine, and apparently to a much smaller extent phosphatidylserine and phosphatidylcholine, present duly in
Gram-negative and Gram-positive bacteria.
6. Significance of Teichoic Acids
The teichoic acid is a polymer invariably found in the wall of certain bacteria. It has
been re-ported that the walls of two
Gram-positive organisms belonging to the genus of micrococci being a member of
the family Micrococcaceae, order Eubacteriales, namely : Staphylococcus aureus, and Staphylococcus faecalis usually comprise
of teichoic acids — i.e., the acidic polymers of ribitol
phos-phate and glycerol phosphate, that
are covalently linked to peptidoglycan, and which can be
conven-iently extracted with cold diluted acids, as given below :
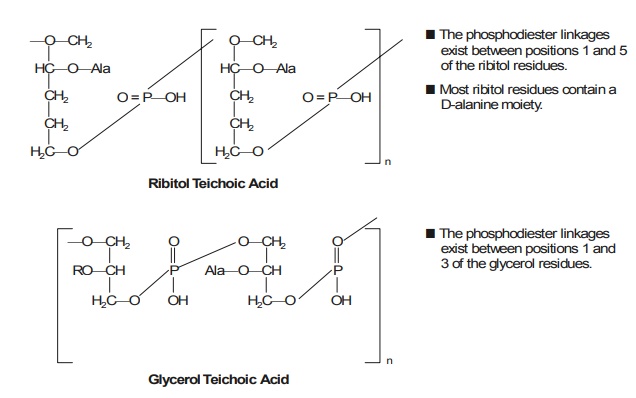
In actual
practice, however, the teichoic* acids may be duly grouped chiefly into two categories, namely : (a) wall
teichoic acids, and (b) membrane teichoic acids.
Characteristic Features : Most teichoic acids do possess certain
inherent characteristic fea-tures as stated here under :
(1) They
usually get bound to Mg2+ ions specifically, and there is quite a
bit of evidence to suggest that they do aid in the protection of bacteria from
the thermal injury by way of
providing an adequate accessible pool of such cations for the stabilization of
the cytoplasmic membrane exclusively.
(2) Importantly,
the walls of a plethora of gram-positive organism contain almost any lipid, but
those which distinctly belong to Mycobacterium,
Corynebacterium, and certain other genera are conspicuously excepted.
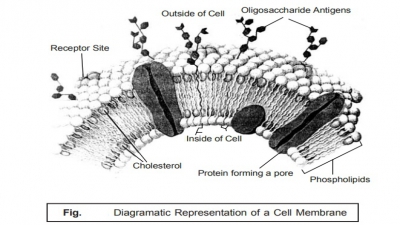
7. The Cell Membrane
Literally,
‘membrane’ designates a thin, soft,
pliable layer of tissue that virtually lines a tube or cavity, covers an organ
or structure, or separates one part from another specifically. The cell membrane refers to the very fine,
soft, and pliable layer of tissue that essentially forms the outer boundary of
a cell ; and it is made of phospholipids,
protein, and cholesterol, with carbohydrates on the outer surface e.g., plasma membrane, as shown in Fig. : 2.14.
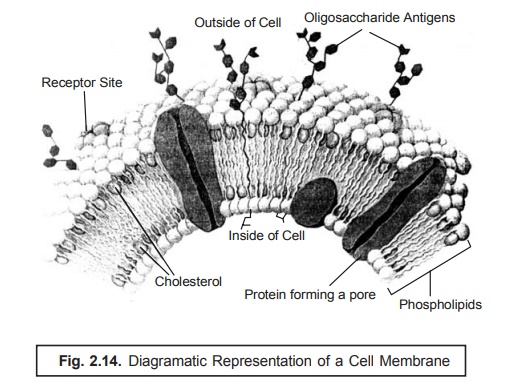
In other
words, the cell membrane is the
bounding layer of the cytoplasmic contents, and repre-sents the principal
osmotic and permeability barrier. It is a lipoprotein (having a ratio of
protein and lipid, 70 : 30), devoid of any polysaccharide, and on being
examined via an electron microscope
shows up with a distinct three-layer unit with a prominent unit membrane
structure.
The
actual thickness of the two outer layers are approximately 3.5 nm, and the
middle layer is nearly 5 nm thick. The lipids observed in the cell membrane are
largely phospholipids, for instance : phosphatidylethanol
amine, and to a lesser extent
phosphatidylserine and . The
other three vital regions in the cell membrane are,
namely :
(a) Polar head regions — of the
phospholipids are strategically positioned at the two outer surfaces,
(b) Centre of membrane —
contain the extended hydrophobic fatty acid chains, and
(c) Middle protein layer — is
duly intercalated into the phospholipid bilayer.
Importance of Cell Membrane. The importance of the cell membrane lies
in monitoring the three vital functions of immense utility
to the cell, namely :
(1) It
mostly acts as an ‘osmotic barrier’,
and usually contains permeases that
are solely respon-sible for the viable transport of nutrients and chemicals
both in and outside the cell ;
(2) It
essentially contains the enzymes that are intimately involved in the
biosynthesis of membrane-lipids together with a host of other macromolecules
belonging to the bacterial cell wall ; and
(3) It
predominently comprises of the various components of the energy generation
system.
It is,
however, pertinent to state here that besides these critically important
features there is an ample evidence to demonstrate and prove that the cell membrane has particular
‘attachment sites’ exclusively meant for the replication and segregation of the
bacterial DNA and the plasmids.
Mesosomes. It has been duly observed that in
certain instances of microorganisms, more specifi-cally and precisely in the Gram-positive bacteria, solely depending
upon the prevailing growth factors as well as parameters the cell membrane
vividly seems to be ‘infolded’ at
more than one point. Such infoldings* are
known as mesosomes as depicted in
Fig. : 2.15.
Habitats. The actual presence of such
folded structures in large quantum have also been found in microorganisms that do possess a relatively higher respiratory
role to play (activity) ;
Examples : (a) Logarithmic phase of
growth, and
(b) Azotobacter i.e., the
nitrogen fixing bacteria.
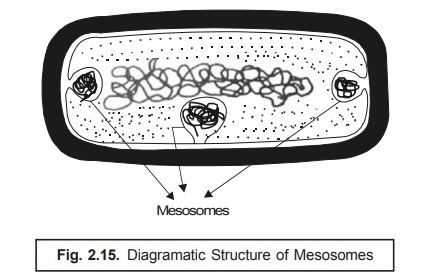
In
addition to the above, the mesosomes
are also found in the following two
types of microor-ganisms, such as :
(i) Sporulating bacteria —
in these the critical appearance of such infolding
(i.e., mesosome formation) is an essential prerequisite for the phenomenon of ‘sporulation’
; and
(ii) Photosynthetic bacteria — in
these the actual prevailing degree of ‘membrane
infolding’ has been intimately related to two important aspects, namely: first — pigment content, and second — photosynthetic
activity.
8. Bacterial Cytoplasm
Based
upon various intensive and extensive investigations carried out on the bacterial cell, one may observe that
the major cytoplasmic contents of it essentially include not only the nucleus
but also ribosomes, proteins, water-soluble components, and reserve material.
It has also been observed that a plethora of bacteria do contain extrachromosomal DNA i.e., DNA that are not connected to the
chromosomes.
It has
also been revealed that the ‘bacterial
nucleus’ is not duly enclosed in a well-defined mem-branous structure, but
at the same time comprises of the genetic material of the bacterial cell.
Interest ingly, several altogether sophisticated meticulous and methodical
investigations pertaining to the actual status/content(s) of the bacterial
nucleus reveal amply that :
(a) Electron microscopy : Electron
micrographs of the bacterial nucleus under investigation evidently depict it as
a region very tightly and intimately packed with fibrillar DNA i.e.,
consisting of very small filamentous structure.
(b) Cytological, biochemical, physical, and genetic
investigations : Such investigations with respect to a large
cross-section of bacterial species revealed that the ‘bacterial nucleus’ essentially contains a distinct singular
molecule of definite circular shape, and having a double-stranded DNA.
The genome size of DNA i.e., the complete set of chromosomes, and thus the entire genetic
information present in a cell, obtained painstakingly from a variety of
bacterial species has been deter-mined and recorded in Table 2.9 below :

Specifications of E. coli: The size of DNA in E. coli together with certain other
specifications are as given below :
Average
length : Approx. 1000 μm
Base
pairs : 5 × 103 kilo base pairs
Molecular
weight : 2.5 × 109 Daltons (± 0.5 × 109)
The
ensuing DNA happens to be a highly charged molecule found to be dissociated
with any basic proteins as could be observed in higher organisms.
Neutralization
of charge is duly caused either by polyamines
e.g., spermine, spermidine, or by bivalent
cations e.g., Mg2+, Ca2+.
Plasmid DNA : Besides, the apparent and
distinct presence of the bacterial ‘nuclear DNA’, they invariably contain
extrachromosomal* DNA termed as plasmid DNA that replicates autonomously.
It has been duly observed these plasmid
DNAs exhibit different specific features, such as :
·
confer on the bacterial cell,
·
drug resistance,
·
ability to generate bacteriocins i.e.,
proteinaceous toxins.
·
ability to catabolize uncommon organic chemical
entities (viz., in Pseudomonas).
Nevertheless,
the actual size of plasmid DNA
usually found in these specific structures may be nearly 1/10th or even less in
comparison to that invariably found in the bacterial
nucleus ; however, the exact number of copies may change from one to
several. Besides, these structures are not enclosed in a membrane structure.
Importantly, the plasmid DNA is
mostly circular in shape and double stranded in its appearance.
9. Ribosomes
Ribosome refers to a cell organelle made
up of ribosomal RNA and protein. Ribosomes may exist singly, in clusters called polyribosomes, or on the surface of rough endoplasmic reticulum. In
protein synthesis, they are the most favoured site of messenger RNA attachment
and amino acid assem-bly in the sequence ordered b the genetic code carried by
mRNA.
In other
words, the specific cytoplasmic area
which is strategically located in the cell material bound by the cytoplasmic
membrane having granular appearance and invariably rich in the macromolecular
RNA-protein bodies is termed as ribosome.
Characteristic Features : Following
are some of the cardinal characteristic features of the ‘ribosomes’, namely:
(1) Contrary
to the animal or plant cells, there exists no endoplasmic reticulum to which ribosomes are bound intimately.
(2) Interestingly,
there are certain ribosomes that are
found to be virtually ‘free’ in the
cyto-plasm ; whereas, there are some, particularly those critically involved in
the synthesis of proteins require to be transported out of the cell, get
closely linked to the inner surface of the cytoplasmic membrane.
(3) The
number of ‘ribosomes’ varies as per
the ensuing ‘rate of protein synthesis’,
and may reach even upto 15,000 per cell. In fact, greater the rate of proteins
synthesis, the greater is the rate of prevailing ribosomes.
(4) Ribosomes represent ribonucleoprotein particles (comprising of 60 RNA ; 40 Protein)
hav-ing a diameter of 200 Å, and are usually characterised by their respective
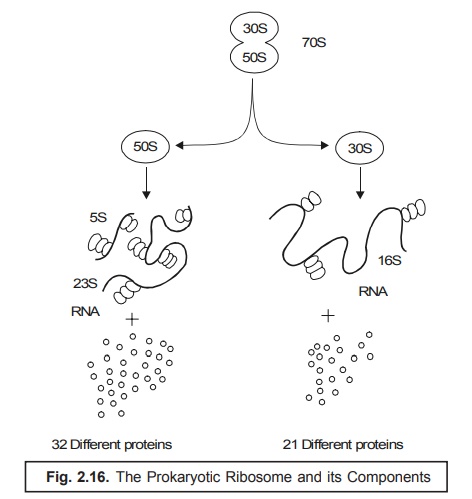
sedimentation physical properties as depicted in Fig. 2.16.
(5) Prokaryotic Ribosome. In the
event when the ribosomes of the prokaryotes undergo ‘sedi-mentation’
in an ultra-centrifuge, they normally exhibit a sedimentation coefficient
of 70 S (S = Svedberg Units), and are
essentially composed of two subunits i.e., a 50 S and a 30 S subunit (almost fused as shown in Figure
2.16). Consequently, these two subunits get dis-tinctly separated into a 50 S
and a 30 S units*. As a result the 50 S unit further gets segre-gated into a
RNA comprised of two daughter subunits of 5 S and 23 S each together with
thirty two (32) altogether different proteins [derived from 50 – (5 + 23) = 22 sub-units].
Likewise,
the 30 S gets fragmented into two segments i.e.,
first, a RNA comprised of only one
subunit having 16 S plus twenty one (21) precisely different proteins [derived
from 30 – 16 = 14 sub-units], (see
Fig. : 2.16).
(6) Eukaryotic Ribosome : This is
absolutely in contrast to the ribosomes of
the corresponding prokaryotic organisms, that do possess a
sedimentation coefficient of 80 S, and are essen-tially comprised of two subunits each of 60 S and 40 S,
respectively.
7. Polysomes. In a situation when these ‘ribosomes’ are specifically
associated with the mRNA in the
course of active protein synthesis, the resulting product is termed as ‘polysomes’.
It is,
however, pertinent to mention here that there are a plethora of ‘antibiotics’ viz., chloramphenicol, erythromycin, gentamycin, and streptomycin,
which exert their predomi-nant action by causing the inhibition of ‘protein synthesis’ in ribosomes.
10. Cellular Reserve Materials
It has
been duly observed that there exist a good number of ‘reserve materials’ strategically located in the prokaryotic cells
and are invariably known as the granular
cytoplasmic inclusions. The three most
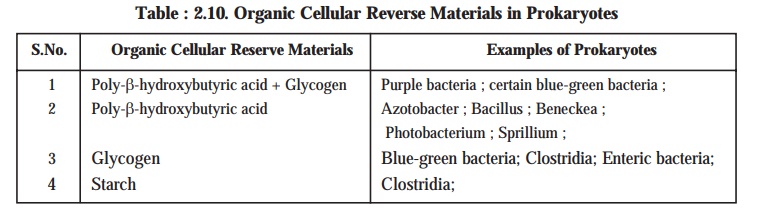
vital and important organic cellular reserve materials present in the prokaryotes are namely : (a) poly-β-hydroxybutyric acid; (b) glycogen; and (c) starch (see Table : 2.10).
Salient Features. The salient features of the organic
cellular reserve materials present in the
prokaryotes are as stated under :
(1) Poly-β-hydroxybutyric
acid. It is found exclusively in the prokaryotes and invariably ca-ters as an equivalent of lipoidal content duly
stored in the eukaryotic cells. It is observed in several species of Azotobacter, bacilli, and pseudomonads.
Interestingly, certain specific or-ganisms viz.,
purple bacteria has the ability to
synthesize even two types of reserve materi-als (e.g., glycogen and poly-β-hydroxybutyrate)
simultaneously.
(a) Visibility — These organic cellular reserve
materials are found to be deposited almost uniformly very much within the
cytoplasm ; however, they may not be detected under a light microscope unless
and until these are duly stained.
(b) Cellular content — The
actively ‘growing cells’ do have
these reserve materials present in rather small quantum in the cellular content
; whereas, they get usually accumulated exclusively in the C-rich culture medium under the influence of restricted amounts of
nitrogen.
(c) Availability — These reserve materials may
sometimes represent even upto 50% of the total cellular content on dry weight
basis.
(d) Utility — These reserve materials are
fully utilized when the prevailing cells are ad-equately provided with a
suitable source of N and the growth is resumed subsequently.
(2) Glycogen and Starch — It has
been duly established that the synthesis of glycogen and starch is usually accomplished via
a proven mechanism for storing C in a form which is osmotically inert ; whereas, in the particular instance of poly-β-hydroxybutyric acid it
pre-cisely designates a method of neutralizing
an acidic metabolite.
(3) Cyanophycine (a copolymer of arginine and
aspartic acid) :
In
general, prokaryotes fail to store
particularly the organic nitrogenous materials, but the blue-green bacteria is expected which essentially accumulate a
nitrogenous reserve material termed
as cyanophycine. It invariably
represents as much as 8% of cellular dry weight; and may be regarded as a
copolymer of arginine and aspartic acid.
(4) Volutin (metachromatic) Granules. A
plethora of prokaryotes acquire more
and more of volutin granules that
may be stained meticulously with a ‘basic dye’, for instance : methyl-ene blue. In fact, these prokaryotes appear as red on being stained with a
‘blue-dye’. Impor-tantly, the prevailing metachromatic nature of the
ensuing ‘red complex’ is on account of the very presence of a substantial
quantum of ‘inorganic phosphates’.
Evidently, the actual accumulation of these substances in the prokaryotes takes
place under critical parameters of starvation specifically during ‘sulphate starvation’. It has been
observed that these instantly generated volutin granules disappear as soon as
the cells are adequately made available with a ‘sulphur source’, and subsequently the phosphate moiety [PO43–] is incorporated
strategi-cally into the nucleic acids i.e.,
DNA and RNA. From the above statement of facts one may vividly infer that the ‘volutin granules’ definitely represent
particularly the ‘intracellular phosphate reserve’ when the desired nucleic
acid synthesis fails to
materialize.
(5) Sulphur Bacteria [e.g., photosynthetic purple sulphur bacteria ; and filamentous
non-photosynthetic bacteria (viz.,
Baggiatoa and Thiothrix)].
The aforementioned two
sulphur bacteria specifically
help in the accumulation of ‘Sulphur’
transiently in the course of hydro-gen sulphide [H2S] oxidation.
(6) Thylakoids. These are solely present in the blue-green bacteria and are intimately
involved in the phenomenon of
photosynthesis. Besides, there are three
prominent structures, namely : gas
vesicles, chlorobium vesicles, and
carboxysomes, that are critically bound by non-unit membranes have been reported to be present in certain photosynthetic organisms.
(7) Ribs. There are several aquatic
prokaryotes essentially
containing gas vacuoles that are intimately
engaged in counter-balancing the prevailing gravitational pull appreciably. On
being examined under a ‘light microscope’ the ensuing gas vacuoles do look like
dense refractile structure having a distinct irregular peripheral
boundary. Importantly, with a cer-tain surge in the hydrostatic built-up
pressure the existing gas vacuoles collapse thereby the cells lose their
buoyancy eventually. Precisely, each gas vesicle more or less has an
appear-ance very much akin to a ‘hollow
cylinder’ having an approximate diameter of 75 nm with distinct conical
ends, and a length ranging between 200 and 1000 nm. These conglomerates of gas
vesicles are usually surrounded by a layer of protein approx. 2 mm thick. These
structures do possess several bands consisting of regular rows of subunits that
almost run perpendicular to the axis, and are termed as ‘ribs’. The ribs are found to be impermeable to water.
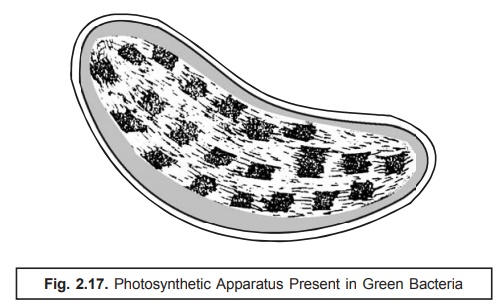
(8) Photosynthetic Apparatus. The photosynthetic apparatus present
specifically in the pho-tosynthetic
green bacteria (chlorobium)
possesses a distinct strategic intracellular loca-tion. It is usually bound by
a series of cigar-shaped vesicles
arranged meticulously in a corticle-layer which immediately underlies the cell
membrane as illustrated in Fig. 2.17. Interestingly, these structures have a
width nearly 50 nm, length varying between 100–150 nm and are delicating
enclosed within a single layered membrane of thickness ranging be-tween 3–5 nm.
They essentially and invariably contain the ‘photosynthetic pigments’.
(9) Carboxysomes. It has
been amply demonstrated that a good number of photosynthetic and chemolithotrophic organisms,
namely : blue-green bacteria, purple bacteria, and thiobacilli essentially
comprise of polyhedral structures having a width of 50–500 nm and carefully
surrounded by a single layer of membrane having a thickness of 3.5 nm
approximately. These characteristic structures are known as carboxysomes. They are found to consist
of certain key enzymes that are closely associated with and intimately involved
in the critical fixation of carbon dioxide [CO2], such as : carboxy dismutase ; and thus, represent
the precise and most probable site of CO2 fixation in the photosynthetic as well as chemolithotrophic organisms.
Related Topics