The Role of the Immune System in Type B Adverse Drug Reactions
| Home | | Pharmacovigilance |Chapter: Pharmacovigilance: Mechanisms of Adverse Drug Reactions
TYPE B OR IDIOSYNCRATIC ADVERSE DRUG REACTIONS : THE ROLE OF THE IMMUNE SYSTEM IN TYPE B ADVERSE DRUG REACTIONS
THE ROLE OF THE IMMUNE SYSTEM IN
TYPE B ADVERSE DRUG REACTIONS
Based
on clinical criteria, it has been postulated that many idiosyncratic ADRs are
immune mediated (Park, Pirmohamed and Kitteringham, 1998; Pirmohamed et al., 1998). Research into this area
is now providing some direct evidence to support the clinical impression. The
mechanism by which a drug leads to an immune-mediated adverse reac-tion is
explained by the hapten hypothesis (Park, Pirmohamed and Kitteringham, 1998)
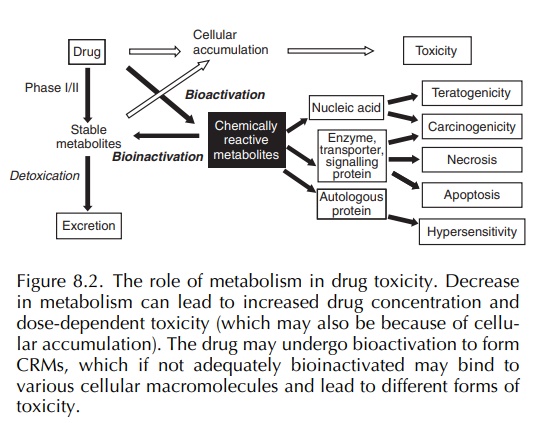
(Figure 8.2). Central to the hapten hypothesis is the assumption that small
molecules such as drugs (<1000 Da) can be recognised as immunogens (i.e. a
substance capa-ble of eliciting a specific immune response) only when they
become covalently bound to an autologous high molecular weight (>50 000 Da)
macromolecular carrier such as a protein (Park, Coleman and Kitter-ingham,
1987). The term ‘hapten’ has been coined to describe such substances that are
not immunogenic per se but become
immunogenic when conjugated to a
macromolecular carrier (this has been termed ‘signal 1’). The type of
hypersensitive reaction will be partly determined by the nature of the immune
response and the site of antigen formation. The best understood reactions are
the type I hypersensitivity reactions induced by penicillins and cephalosporins
and mediated by immunoglobulin E (IgE) antibod-ies directed against a drug
hapten conjugated to protein (Pirmohamed, Kitteringham and Park; Weiss and
Adkinson, 1988). Severe anaphylactic reactions occur in only few patients (1 in
2000); atopic patients are at increased risk, although the genetic basis of
this and of the IgE response to penicillins remains to be elucidated.
Less
well understood are the immunological mech-anisms that underlie severe skin
reactions such as Stevens–Johnson syndrome (SJS) and immuno-allergic hepatitis.
There is clear chemical evidence from in
vitro studies that the drugs associated with these reactions can undergo
oxidative metabolism to CRMs that can haptenate proteins (Park, Pirmohamed and
Kitteringham, 1995). In addition, both humoral and cell-mediated responses
directed against drug-induced antigen have been detected in patients, e.g. in
halothane hepatitis (Pohl et al.,
1990). With some compounds, the immune response seems to be directed
predominantly towards an auto-antigen. For example, in tienilic acid-induced
hepatitis, patients have circulating auto-antibodies directed against the P450
isoform (CYP2C9), which is responsible for the bioactivation of tienilic acid
(Beaune and Bourdi, 1993).
The
fundamental concept that protein conjugation is an obligatory step in the
process of immune recog-nition of drugs has however recently been chal-lenged
by the observation that T-cell clones from patients hypersensitive to many
drugs undergo prolif-eration in an antigen-processing–independent [but major
histocompatibility complex (MHC)-restricted] manner (Schnyder et al., 1997; Zanni et al., 1998). This requires labile, reversible binding of drug to
the MHC complexes on antigen-presenting cells. The presence of T-cell clones
that proliferate only in response to parent drug rather than metabolite and the
rapid down-regulation in expression of the T-cell receptor upon stimulation are
consistent with this mechanism. It is of course possible that both mech-anisms
may be important in the overall pathogene-sis. For example, the
hapten-dependent pathway may be more important for primary immune stimulation
(sensitization), whereas the metabolism-independent pathway may be all that is
necessary for secondary stimulation and elicitation of tissue damage
(Pirmo-hamed, Naisbitt and Park, 2001). Further studies are needed to define the
roles of the two pathways of drug (antigen) presentation in the pathogenesis of
immune-mediated ADRs.
Irrespective
of the mechanism of antigen presen-tation, T cells are of fundamental
importance in the immune response against a drug (Naisbitt et al., 2000a). The interaction between the T cell and the drug
(antigen) in the groove of the MHC governs the immune response. MHC class I
molecules bind peptides of 8–10 amino acids and present to CD8+ T cells (Pamer and Cresswell,
1998). MHC class II molecules present longer peptide molecules (13–17 amino
acids) to CD4+ cells (Jensen,
1997). Although class I molecules are found on all cell surfaces, class II
molecules are only expressed on specialised antigen-presenting cells such as
macrophages but can become expressed on other cells such as keratinocytes in
the presence of pro-inflammatory cytokines such as interferon-gamma (INF- )
(Pichler and Yawalkar, 2000). The nature of the immune response is governed by
the differentiation of T cells into T-helper 1 TH1 TH2,
T-cytotoxic 1 TC1 or TC2 subsets. TH1 and TC1
cells mediate cytotoxicity and local inflammatory reactions, whereas TH2
and TC2 cells stimulate B-cell-dependent antibody production
(Romagnani, 1999).
It
is important to note that the presence of an anti-gen (i.e. signal 1) in the
absence of co-stimulatory molecules will lead to tolerance and T-cell apoptosis
(Naisbitt et al., 2000a). Although
the role of surface molecules such as B7.1 and B7.2 as co-stimulatory molecules
has long been known, the importance of cytokines has only been recognised
recently. In addi-tion to signal 1, two other signals are required to stimulate
a full immune response (Curtsinger et al.,
1999). Signal 2 is represented by a series of pro-inflammatory cytokines such
as interleukin-2 (IL-2), TNF- and IFN- that act indirectly on
antigen-presenting cells to up-regulate the expression of co-stimulatory
molecules. Signal 3 represents polarising cytokines that act directly on T
cells. It is known that TH1 cells produce IL-12 and IFN- , which
promote the activation of macrophages and cell-mediated immunity. By contrast,
TH2 cells produce IL-4 and IL-13; these provide help for the humoral
immune response by promoting IgG to IgE class switching.
An
interesting hypothesis termed the ‘danger hypothesis’ has recently been
proposed in the field of immunology to explain the basis of self-tolerance
(Anderson and Matzinger, 2000; Gallucci and Matzinger, 2001; Matzinger, 1994).
This can also be applied to the mechanism of drug hypersensitivity (Park, Pirmohamed
and Kitteringham, 1998; Uetrecht, 1999). This hypothesis states that the immune
system responds to most antigens with tolerance, and only in the presence of a
danger signal will the presentation of an antigen result in an immune response.
The nature of the danger signals has not been accurately defined, but
pro-inflammatory and polarising cytokines, intracel-lular contents resulting
from cell necrosis and exoge-nous proteins including those derived from
viruses, are all potential candidates (Gallucci and Matzinger, 2001). With
respect to drug hypersensitivity, it can be hypothesised that the CRM may not
only provide signal 1 (by conjugating with a protein) but also provide the
co-stimulatory signals 2 and 3 by the activation of signalling pathways linked
to oxidative stress and protein damage, including the secretion of cytokines
(Park et al., 2001). Furthermore, the
hypoth-esis also allows the possibility that the co-stimulatory molecules are
completely independent of the drug and could be, for example, concomitant viral
infections (see THE ROLE OF VIRUSES IN
TYPE B ADVERSE DRUG REACTIONS).
Related Topics
