Bacterial Transduction
| Home | | Pharmaceutical Microbiology | | Pharmaceutical Microbiology |Chapter: Pharmaceutical Microbiology : Microbial Genetics and Variations
Bacterial transduction may be defined as - ‘a phenomenon causing genetic recombination in bacteria wherein DNA is carried from one specific bacterium to another by a bacteriophage’.
Bacterial
Transduction
Bacterial transduction may be
defined as — ‘a phenomenon causing
genetic recombination in bacteria wherein DNA is carried from one specific
bacterium to another by a bacteriophage’.
It has
been duly observed that a major quantum of bacteriophages, particularly the ‘virulent’ ones, predominantly undergo
a rather quick lytic growth cycle in
their respective host cells. During this phenomenon they invariably inject
their nucleic acid, normally DNA, right into the bacterium, where it takes up
the following two cardinal steps :
(a) undergoes
‘replication’ very fast, and
(b) directs
the critical synthesis of new phage
proteins.
Another
school of thought may put forward another definition of bacterial transduction as - ‘the
actual and legitimate transfer by a bacteriophage, serving as a vector, of a
segment of DNA from one bacterium (a donor) to another (a recipient)’.
Zinder
and Ledenberg (1952) first and foremost discovered the wonderful phenomenon
during an intensive search for ‘sexual
conjugations’ specifically amongst the Salmonella
species.
Methodology :
The various steps that are involved in the bacterial transduction phenomenon are as stated under :
(1) Auxotrophic
mutants were carefully mixed together ; and subsequently, isolated the
prototrophic recombinant colonies from the ensuing selective nutritional media.
(2) U-Tube Experiment : The
U-tube experiment was duly performed
with a parental auxotrophic strain
in each arm (viz., I and II), and
adequately separated by a microporous
fritted glass (MFG) filter, whereby the
resulting ‘prototrophs’
distinctly appeard in one arm of the tube, as shown in Fig. 6.10.
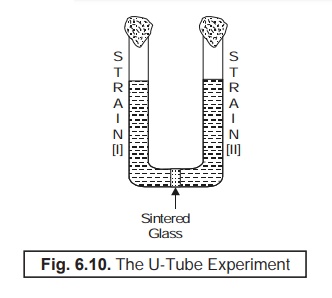
As the
MFG-filter particularly checked and prevented cell-to-cell contact, but at the
same time duly permits the ‘free
pas-sage’ of fluid between the said two
cultures [i.e., strains I and II], it may be safely inferred that there must
be certain ‘phenomenon’ other than
the ‘conjugation’ was involved.
Besides,
the process could not be radically prevented to DNAase (an enzyme) activity,
thereby completely eliminating ‘transformation’
as the possible phenomenon involved for causing definite alterations in the
recipient auxotrophs to prototrophs.
The
bacteriophage was duly released in a substantial amount from a lysogenic (i.e., recipient) culture. Thus, the emerging phage
critically passed via
the MFG-filter, and adequately
infected the other strain (i.e., donor) lyzing it exclusively.
Finally,
during the ‘replication’ observed in
the donor strain, the ensuing phage
adventitiously comprised of the relevant portions of the critical bacterial chromosome along with it. Eventually, it gained
entry via the MFG-filter once again ; thereby taking with it a certain viable
segment of the respective donor’s ‘genetic
information’ and ultimately imparting the same to the desired recipient strain.
Nevertheless,
the ‘bacterial transduction’ may be
further classified into two
sub-heads, namely :
(a) Generalized
transduction, and
(b) Specialized
transduction.
(a) Generalized Transduction
In a
situation when practically most of the fragments pertaining to the bacterial DNA* do get an obvious chance
to gain entry right into a ‘transducting
phage’, the phenomenon is usually termed as ‘generalized transduction’.
Modus
Operandi : The very first step of the phage
commences duly with the ‘lytic cycle’ whereby the prevailing ‘viral enzymes’ preferentially hydrolyze the specific bacterial chromosome essentially into several small fragments of DNA. In fact, one
may most conveniently incorporate any portion of the ‘bacterial chromosome’ right into the ‘phage head’ in the course of the ensuing phage assembly ; and, therefore, it is not normally associated with
any sort of ‘viral DNA’.
Example :
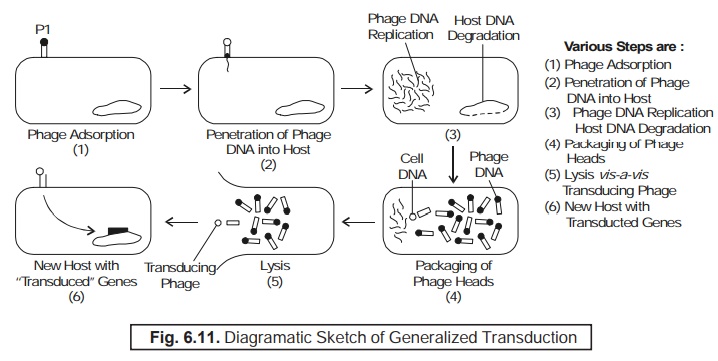
Transduction of Coliphage P1 : In fact,
the coliphage P1 can effectively transduce a variety of genes in the bacterial chromosome**. After
infection a small quantum of the phages carry exclusively the bacterial
DNA as shown in Fig. 6.11.
Figure :
6.11 clearly illustrates the following salient
features, namely :
Phage P1 chromosomes, after
injection into the host cell, gives rise to distinct degradation of the specific host chromosome right
into small fragments.
During
maturation of different particles, a small quantum of ‘phage heads’ may, in fact, envelop certain fragments of the bacterial DNA instead of the phage DNA.
Resulting
bacterial DNA on being introduced
into a new host cell may get integrated into the bacterial chromosome, thereby causing the transference of several genes from one host cell to another.
It has
been observed that the ‘frequency’
of such defective phage particles
usually range between 10– 5 to 10– 7 with respect to
corresponding ‘progeny phage’
generated. As this particular DNA more or less matches the DNA of the newer
bacterium thus infected, the ‘recipient
bacterium’ shall not be rendered lysogenic*
for the respective P1 phage.
Instead, the injected DNA shall be duly integrated right into the chromosome of
the available recipient cell. In this manner, the so called ‘genetic markers’ duly present in the DNA would precisely detect the very
presence of all defective P1 phages essentially
bearing the E. coli DNA.
Advantages :
The various glaring advantages of
the generalized transduction are as
given below :
(1) Just
like bacterial conjugation (see
Section 2.10) and bacterial
transformation (see Sec-tion 2.7) the generalized
transduction also caters for the typical ways for ‘mapping* bac-terial genes’, by virtue of the fact that the
fragments duly transferred by the bacteriophage are invariably big enough to safely accomodate hundreds of genes.
(2) To
test actually the exact quantum of such ‘recombinants’
that have inherited from other ‘donor
markers’ due to the growth occurring on other culture media.
(3) Strategic
closeness of the ‘two markers’ on
the bacterial chromosome ascertains
the fact that they would be inherited together more likely by the aid of a single transducing phage.
(b) Specialized Transduction
Based on
enough scientific evidences it has been duly proved and established that the ‘bacterial genes’ may also be adequately transduced by means of bacteriophage
in another equally interesting and thought
provoking phenomenon usually termed as ‘specialized
transduction’. In fact, this phenomenon confirms duly that certain template phage strains may be capable
of transferring merely a handful of ‘restricted
genes’ belonging categorically to the
‘bacterial chromosomes’.
In other
words, the ensuing phages particularly transduce exclusively such bacterial genes that are strategically
positioned quite adjacent to the prophage
in the bacterial chromosome.
Therefore, this particular process is sometimes also referred to as ‘restricted transduction’.
Interestingly, in an event when such a phage duly infects a cell, it invariably
carries along with it the specified group
of bacterial genes which
ultimately turns out to be an integral
part of it. Consequently, such genes may recombine meticulously with the homologous DNA of the prevailing infected cell.
Phage Lambda (λ) of E. coli. : In a
broader perspective, the most elaboratedly researched spe-cialized transducting phage is duly represented by the phage lambda (λ) of E. coli.
The exact
location of the ensuing λ prophage present in the bacterial chromosome invariably lies
between the bacterial genes gal
and bio. It may be observed that whenever phages duly carrying
either a gal or bio genes do infect an altogether ‘new
host’, then the desired recombination either with the gal or bio
genes of the respective may take place articulately. Fig. 6.12 depicts vividly
the phenomenon of specialized transduction.
Salient Features :
The
various salient features highlighting
the process of specialized transduction in
Figure 6.12 are stated as under :
(1) Practically
‘all phages’ which essentially carry certain bacterial genes solely on account of ‘‘incorrect’’ excision are obviously found to the ‘defective’ with respect to certain highly important functions.*
(2) Thorough
passage via the entire ‘replication cycle’ cannot be accomplsihed
; whereas, the ensuing cell may suitably give rise to certain phages, provided it is also duly infected with a rather ‘complete phage’.
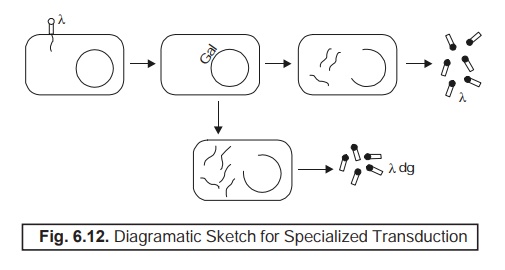
Explanations :
The various stages illustrated in
Fig. 6.12 are as follows :
(1) When
a cell gets duly infected by phage λ, its DNA is precisely inserted
right into the bacterial genome next
to the genes meant for galactose
metabolism (i.e, gal genes).
(2) Invariably
when such a cell is being induced, the λ DNA
emerges out promptly, get repli-cated, and subsequently turned into a normal phage.
(3) Sometimes,
the respective λ DNA is
excised imperfectly thereby taking along with it the gal genes ; and hence
leaving behind certain quantum of itself that may finally lead to λ dg (i.e., defective-galactose transducing phage.)
Related Topics
