Erythrocytes
| Home | | Anatomy and Physiology | | Anatomy and Physiology Health Education (APHE) |Chapter: Anatomy and Physiology for Health Professionals: Blood
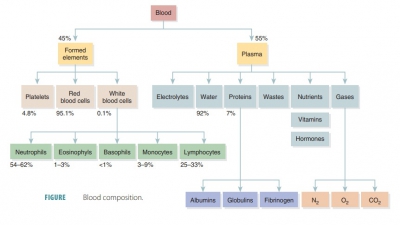
The formed elements of blood include erythrocytes, leukocytes, and platelets.
Formed Elements
The formed elements of blood include erythrocytes,
leukocytes, and platelets. Red blood cells make up about 45% of blood volume,
which is known as the hematocrit.
The RBCs have no nuclei or organelles,meaning that they are not considered
“true cells.” Platelets are only cell fragments. Only leukocytes are complete
cells. Leukocytes and platelets make up less than 1%. The remainder is plasma.
Most blood cells do not divide. Instead they are replaced when stem cells continuously
divide in the bone marrow. All formed elements arise from the hematocytoblasts, also called hematopoietic stem cells, which are undifferen-tiated precursor
cells in the red bone marrow. Most formed elements only survive in the
bloodstream for a few days. Different modes of maturation of formed elements
exist. Once a cell becomes committed to a certain blood cell pathway, it is unable
to change. Membrane surface receptors appear, which signal the cell’s
commitment to one blood cell pathway. The receptors respond to specific growth
factors or hor-mones. These assist the cell in becoming even more specialized.
In a healthy male adult the normal hema-tocrit value is 47%, plus or minus 5%.
In a healthy female adult, it is 42%, plus or minus 5%. Less than 1% of blood
volume consists of platelets and leukocytes. Most of the remaining 55% of whole
blood is made up by the plasma (FIGURE
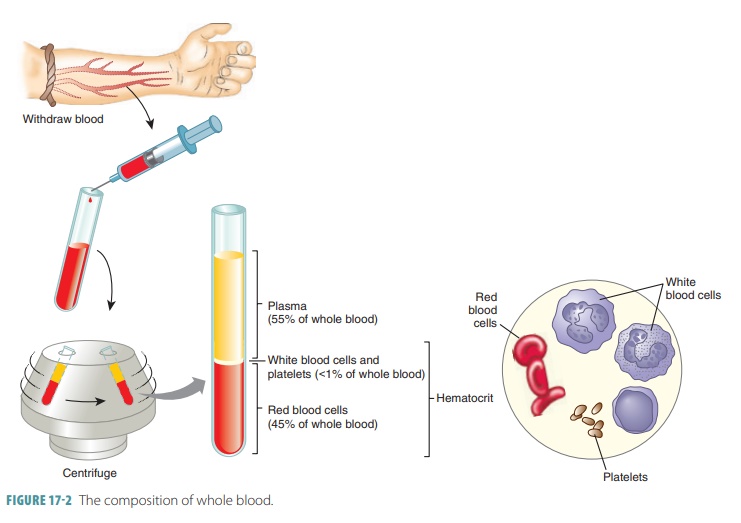
17- 2). The components that make up whole blood can be separated
or fractionated to be clinically
analyzed.
Erythrocytes
Erythrocytes
or red
blood cells (RBCs) have abiconcave shape, meaning they are
basically round, with a center that is depressed in comparison withtheir
edges. This shape helps them to transport gases by increasing the surface area
of the cell, allow-ing more diffusion. This shape also ensures that the cell
membrane is nearer to the hemoglobin, which
carries oxygen, inside the cell. Erythrocytes, when mature, are bound by a
plasma membrane. Red blood cells are about one-third hemoglobin, a protein that
gives them their red color. Therefore, hemoglobin is the major protein in red
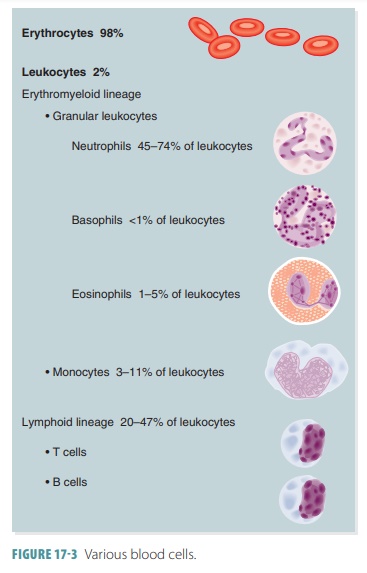
blood cells. FIGURE
17-3 shows the various types of blood cells.
The formation of erythrocytes, called erythropoiesis, occurs only in the red bone mar-row or the myeloid tissue, the tissue that performs hematopoiesis. Erythropoiesis increases whenoxygen levels in the blood decrease. Erythrocytes have nuclei that are shed as they mature, allowing more room for hemoglobin. Lacking nuclei, mature RBCs cannot synthesize proteins or divide to form more cells. They produce adenosine triphosphate through glycolysis because they do not have mito-chondria and use none of the oxygen carried in their hemoglobin. Erythrocytes also have nearly no organelles and contain mostly antioxidant enzymes and structural proteins. The structural proteins allow them to change shape and return to their orig-inal shape afterward. A network of proteins, primar-ily one called spectrin, is attached to the cytoplasm of red blood cell plasma membranes. This maintains the biconcave shape. Spectrin forms a net that allows RBCs to bend, turn, and become more concave as needed to move through tiny capillaries. For exam-ple, in lung capillaries RBCs pick up oxygen and release it to tissue cells through other body capillar-ies. They also move approximately 20% of the car-bon dioxide from tissue cells back to the lungs. Red blood cells are highly efficient in these tasks because they generate adenosine triphosphate via anaerobic mechanisms and do not have mitochondria. This means they do not consume any of the oxygen they carry. Oxygen is picked up from the alveoli by the red blood cells, and it binds with hemoglobin. The RBCs carry oxygen to the tissue cells. They can form stacks called rouleaux.
Hemoglobin
Hemoglobin is responsible for the ability of the cells to
transport oxygen and carbon dioxide, and most oxy-gen carried in the blood is
bound to hemoglobin. It also aids in the process of blood clotting. Hemoglo-bin
consists of the red heme pigment
that is bound to the globin protein.
Hemoglobin makes up more than 95% of the protein in a red blood cell. Globin
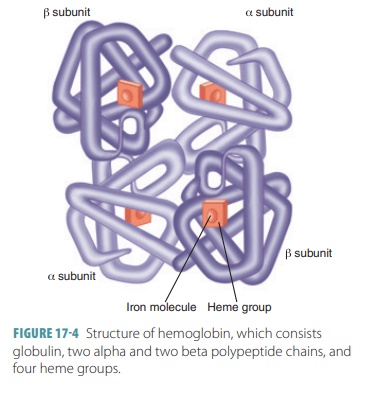
has four polypeptide chains (two alpha and two beta) that each bind a ring-like
heme group (FIGURE
17-4). Each heme group has an iron atom in its very center. One
hemoglobin molecule can transport four molecules of oxygen, since each iron
atom combines reversibly with one molecule of oxygen. One red blood cell
consists of about 250 million hemoglobin molecules, meaning that each of them
can contain approximately one bil-lion oxygen molecules. Because hemoglobin is
inside erythrocytes, it does not break into fragments that could leak out of
the bloodstream through capillary walls. The containment of hemoglobin by the
eryth-rocytes also keeps it from making the blood more vis-cous and raising
osmotic pressure.
When hemoglobin easily and reversibly binds with oxygen,
oxyhemoglobin is formed. Oxyhemoglobin is bright
red and has a three-dimensional structure. When oxygen is released,
deoxyhemoglobin is formed. Deoxyhemoglobin
is burgundy (darker red), andblood rich in deoxyhemoglobin may
appear bluish when seen through blood vessels. Deoxyhemoglobin is also known as
reduced hemoglobin.
Approximately 20% of the carbon dioxide that is transported
by the blood is combined with hemoglobin. However, the carbon dioxide binds to
the amino acids of the globin portion of hemo-globin instead of the heme portion. This forms carbaminohemoglobin, and the process
occursmore easily when the hemoglobin is dissociated from oxygen. This is known
as its reduced state . The loading of
carbon dioxide occurs in the tissues, with transport occurring from the tissues
to the lungs, where it is eliminated from the body.
Mature RBCs contain an adult-type of hemoglo-bin called HbA.
In an embryo or fetus, a different form of hemoglobin, known as fetal hemoglobin or HbF, is contained in the RBCs. This binds oxygen more readily than
adult hemoglobin. Therefore, a developing fetus can “steal” oxygen from the
mother’s bloodstream via the placenta. Fetal hemoglobin begins to convert to
adult hemoglobin shortly before birth and continues over the next year.
The way hemoglobin is contained in the RBCs prevents two
major occurrences. The hemoglo-bin does not break into fragments and therefore
does not leak out of the bloodstream through the walls of the capillaries.
Also, the hemoglobin is prevented from increased blood viscosity and rais-ing
osmotic pressure. The viscosity of the blood is mostly determined by the
erythrocytes. When the number of RBCs exceeds the normal range, blood flows
more slowly because it has become more vis-cous. Oppositely, blood becomes
thinner and flows more quickly when the RBC count drops below the normal range.
A
red blood cell count is the number of RBCs in a microliter of blood. Normal
ranges are as follows:
■■
Adult males: 4.7 million to 6.1 million cells per microliter
■■
Adult females: 4.2 million to 5.4
million cells per microliter Increased numbers of circulating RBCs increase the
blood’s oxygen-carrying capacity, which can affect health positively. Red blood
cell counts are taken to diagnose many diseases and evaluate their courses.
Erythropoiesis
In humans, RBCs are mostly developed in spaces within bones
that are filled with red bone marrow. Erythrocytes usually live for 120 days,
with replace-ment cells created to maintain a relatively stable RBC count. The
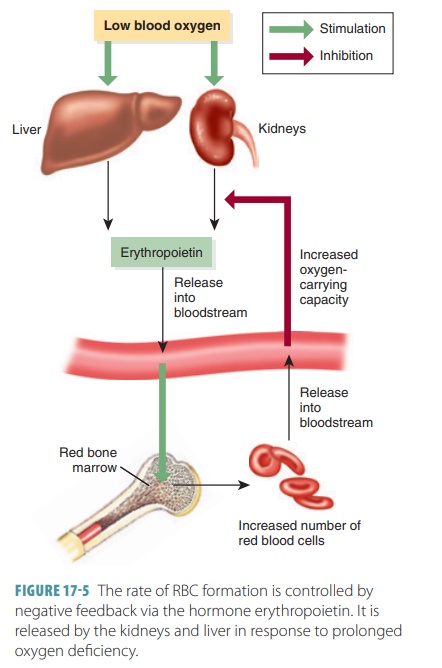
rate of RBC formation is controlled by neg-ative feedback via the hormone erythropoietin. It is released by the kidneys and
liver in response to pro-longed oxygen deficiency (FIGURE 17-5).
The formation of all types of blood cells is known as hematopoiesis and occurs in the red bone mar-row. In this bone marrow, there is a soft network of reticular connective tissue. This tissue bordersbloodsinusoids, which are wide blood capillaries. The net-work contains immature blood cells, fat cells, mac-rophages, and reticular cells. Blood reticulocyte counts provide information regarding the rate of erythrocyte formation. The reticular cells secrete the connective tissue fibers. In an adult, red bone mar-row is mostly found in the axial skeleton bones and girdles and in the proximal epiphyses of the femur and humerus.
Production of RBCs continues at a heightened rate until the amount of them in the blood circulation is enough to supply oxygen to the body tissues. The stages of formation of RBCs and other types of blood cells from hemocytoblasts are shown in FIGURE 17-6. These stages have been individually named by hema-tologists. Approximately 1 ounce of new blood is created every day. This ounce of blood contains approximately 100 billion new cells.
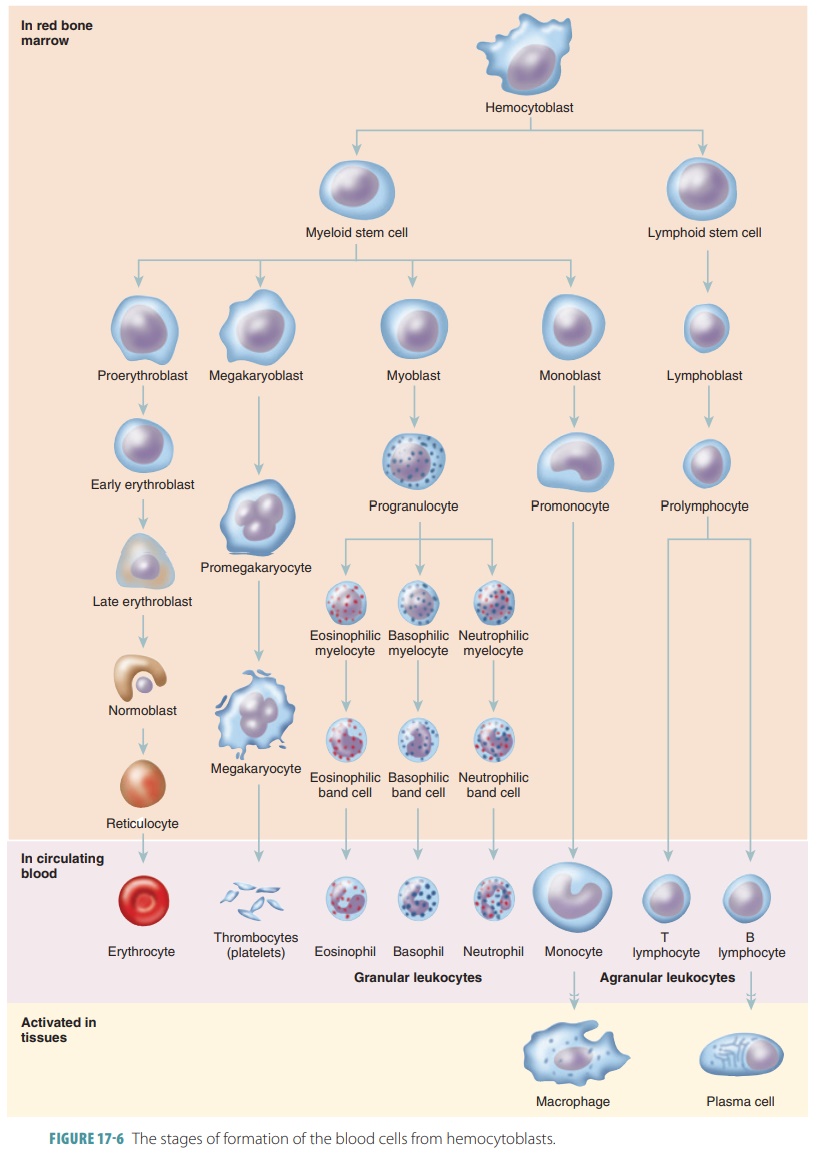
The stages of RBC maturation are described briefly, as
follows:
■■ Hemocytoblasts or multipotent stem cells, in the red bone
marrow, produce myeloid
stem cells; myeloid stem cell division creates progenitor cells, from which all formed elements derive, except for
lymphocytes.
■■ The myeloid stem cells divide,
producing RBCs and several types of WBCs.
■■ Lymphoid
stem cells divide to produce varioustypes of lymphocytes.
■■ Cells that will become RBCs initially differentiate into
proerythroblasts.
■■ These cells then mature through several erythro blast
stages, actively synthesizing hemoglobin.
■■ After approximately four days, the erythroblasts are then
called normoblasts and shed their nuclei to become reticulocytes; the
reticulocytes contain 80% of the hemoglobin of a mature RBC and are immature
cells found in the peripheral blood.
■■ Hemoglobin synthesis continues for up to three more days;
the cytoplasm of these cells still contains RNA.
■■ The reticulocytes move from the bone marrow into the
bloodstream.
■■ Twenty-four hours later, the reticulocytes are fully
matured and cannot be distinguished from other mature RBCs.
B-complex vitamins such as vitamin B12 and folic
acid greatly influence RBC production and are nec-essary for DNA synthesis.
Hematopoietic or bloodcell–forming tissue
is very vulnerable to deficiency ofboth of these vitamins. Iron is required for
normal RBC production and for hemoglobin synthesis. Iron is slowly absorbed
from the small intestine, and the body reuses much of the iron released by
decomposi-tion of hemoglobin from damaged RBCs. Only small amounts of iron must
be taken in via the diet.
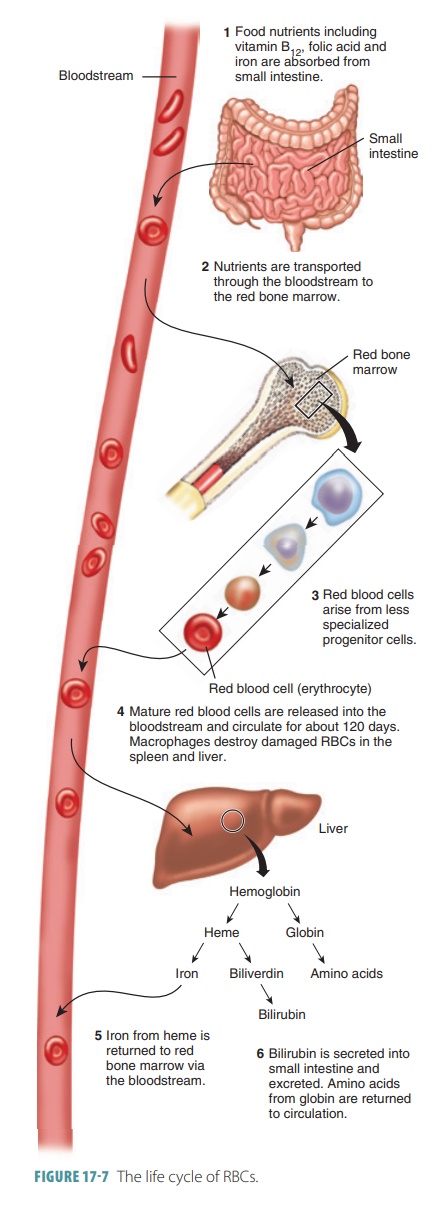
Breakdown of Erythrocytes
Red blood cells bend as they move through blood vessels, but
aging causes them to become more fragile. Cells called macrophages phagocytize and destroy damaged
RBCs, mostly in the liver and spleen. Hemoglobin from RBCs is broken down into heme, which contains iron, and the
protein globin.The heme then
decomposes into iron and biliverdin, a green
pigment. The iron may be transported by the blood to synthesize new
hemoglobin, with about 80% of the iron stored in the liver as an iron–protein
complex. Biliverdin is converted into bilirubin, an orange pigment that is excreted along with biliverdin
in the bile. The life cycle of RBCs is summarized in FIGURE 17-7.
Erythropoietin Regulation
Especially during hypoxia, the kidneys produce erythropoietin,
which is also called erythropoiesis-
stimulating hormone. Erythropoietin stimulates pro-duction of erythroblasts
from bone marrow. Therefore, bone marrow can increase the rate of RBC formation
by approximately 10 times—about 30 million cells per second. This process aids
the patient during recovery from a severe loss of blood.
Erythropoietin
is
actually a glycoprotein hormone.If certain kidney cells are deficient in
oxygen, enzymes that are oxygen-sensitive cannot function normally. Hypoxia-inducible factor is an
intracellular signalingmolecule that accumulates as a result, increasing the
synthesis and release of erythropoietin. This condi-tion may be caused by iron
deficiency, in which there is insufficient hemoglobin in each RBC; high
altitudes or pneumonia, which cause reduced availability of oxygen; excessive
destruction of RBCs; or hemor-rhage, which results in reduced numbers of RBCs.
Oppositely, erythropoietin production is depressed by excessive oxygen or
excessive erythrocytes in the bloodstream. In the bloodstream, erythropoietin
stimulates red bone marrow cells that are already committed to forming
erythrocytes, causing them to mature more quickly.
Iron
Amino acids, lipids, and carbohydrates are required for
erythropoiesis to occur. Hemoglobin synthesis requires iron, which is available
in the diet. The cells of the intestines accurately control iron absorption
into the bloodstream as a result of changing amounts of iron that are stored in
the body. Hemoglobin con-tains approximately 65% of the body’s supply of iron,
which is about 4,000 mg. The liver and spleen store most of the rest of body
iron, with small amounts also stored in the bone marrow. Iron is stored inside cells
as protein–iron complexes called ferritin and hemo-siderin,because
free iron ions are toxic to the body.
Iron is loosely bound to a transport protein called a metalloprotein, which transports globulin. Erythro-cytes
that are developing take up iron as required to form hemoglobin. Every day we
lose small amounts of iron via the feces, perspiration, and urine. Men lose
approximately 0.9 mg per day, and women lose approximately 1.7 mg per day.
Menstruation is the rea-son for additional average daily loss of iron in women.
As erythrocytes age they become less flexible and more
fragile and rigid. The hemoglobin degenerates, with these RBCs fragmenting and
becoming trapped in the smaller vessels, mostly those of the spleen. They are
engulfed and destroyed by macrophages, with the heme portion of their
hemoglobin being split from the globin portion. The iron core is saved and
bound to ferritin or hemosiderin for future use.
1. Describe the function of hemoglobin.
2. Explain how RBCs produce adenosine triphosphate.
3. Which hormone is required for the formation of
erythrocytes?