Active Cell Mechanisms
| Home | | Anatomy and Physiology | | Anatomy and Physiology Health Education (APHE) |Chapter: Anatomy and Physiology for Health Professionals: Levels of Organization : Cells
Active cell mechanisms include active transport, endocytosis, and exocytosis.
Movements Through Cell Membranes
The cell membrane controls the substances that can enter and leave the cell. It does this by using passive and active mechanisms. Passive mechanisms do not require cellular energy, whereas active mechanisms do.
Active Cell
Mechanisms
Particles sometimes move from a
region of lower concentration to a region of higher concentration. When this
occurs, energy is required. This energy comes from the cellular metabolism,
specifically from the molecule known as ATP, which is created in the
mitochondria of cells. Active cell mechanisms include active transport,
endocytosis, and exocytosis.
Active Transport
Active
transport is the movement of
particlesthrough membranes from regions of lower concen-tration to regions of
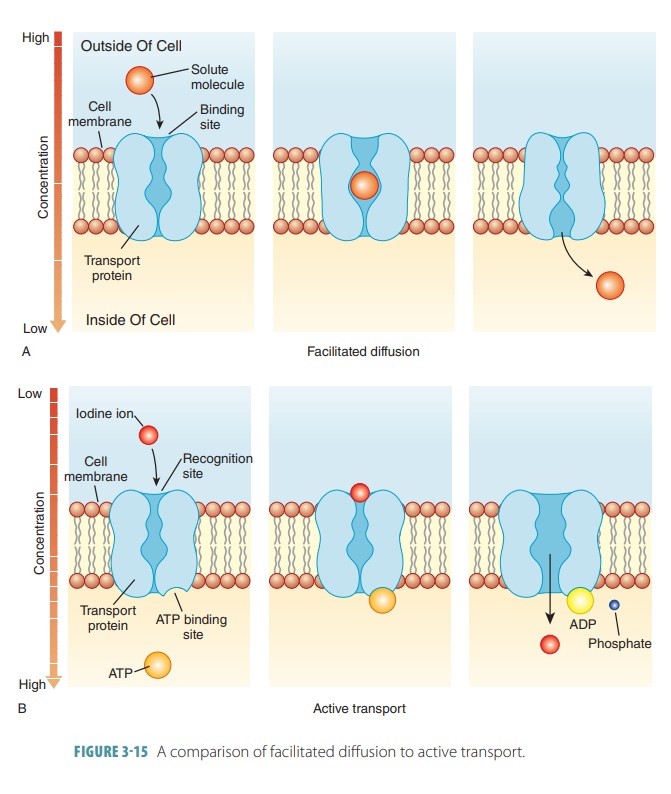
higher concentration. Similar to carrier-mediated facilitated diffusion, it
also requires carrier proteins, which combine with transported substances both
specifically and reversibly. This pro-cess is similar to facilitated diffusion
because of its use of specific carrier molecules in
the cell membranes (FIGURE 3 -15). These carrier molecules are proteins that have binding sites that
combine with the parti-cles they are carrying. However, active transport
dif-fers from facilitated diffusion in that ATP is required. Active transport
moves sugar, amino acid, sodium, potassium, calcium, and hydrogen particles
across cell membranes. Active transporters or solute
pumps move solutes, mostly ions, against the concentration gradient, requiring
energy. There are two types of active transport: primary and secondary.
Primary Active Transport
In primary
active transport, required energy comes directly
from the hydrolysis of ATP. Hydrolyzed ATP causes phosphorylation of the
transport protein, chang-ing its shape so it pumps the bound solute across the
membrane. Calcium and hydrogen pumps are primary active transport systems.
However, perhaps the best studied of these systems is the sodium-potassium pump, which uses the enzyme sodium-potassium ATPase (Na+-K+ ATPase).
Remember that the concentration of potassium inside cells is much higher than
outside (by about 10 times), and the reverse is true of sodium. These balances
are used for essential muscle and nerve cell function and to maintain normal
fluid volume inall body cells. Sodium and potassium leak continu-ously yet
slowly through leakage channels in the plasma membrane. They cross more quickly
in stimulated mus-cle and nerve cells. Therefore, the sodium-potassium pump
acts as a nearly continuous antiporter. It drives sodium out of cells, against
a large concentration gradi-ent, as it pumps potassium back into them.
Ions driven by a concentration
gradient may be slowed in their movement by the negative or positive charges of
certain plasma membranes. Ions realisti-cally diffuse according to electrochemical gradients. Therefore, these gradients used by the sodium- potassium pump are the
basis for most secondary active transport of ions as well as nutrients, of
vital importance to cardiac, neuronal, and skeletal muscle function.
Secondary Active Transport
In the secondary active transport, the process indi-rectly uses energy that is stored in ionic gradients.
These are created by primary active transport pumps. Second-ary active
transport is a coupled system that
moves more than one substance at a time. There are two subforms: in a symport system, the two substances are
transported in the same direction, and in an antiport system, they cross the membrane in opposite directions.
One ATP-powered pump can
indirectly drive the secondary active
transport of a few other solutes. The pump stores energy in the ion
gradient by mov-ing sodium against its concentration gradient, across the
plasma membrane. A substance pumped across a membrane can accomplish work as it
leaks back down along its concentration gradient. Therefore, other sub-stances
are cotransported as sodium moves back into the cell via carrier proteins (a
symport system). An example is the secondary active transport of certain
sugars, amino acids, and ions into the cells that line the small intestine.
Because the concentration gradient of the ion is used for energy, the ion has
to be pumped out of the cell in order to maintain its diffusion gradient.
Antiport systems can also be
driven by ion gradi-ents. An example of such an antiport system is one that
helps regulate intracellular pH by using the sodium gradient for the expulsion
of hydrogen ions. Each membrane pump or cotransporter transports only specific
substances, no matter how energy is acquired to do so. When substances cannot
pass by diffusion, the cell uses active transport systems to be selective. If
there is no pump, nothing can be transported.
Vesicular Transport
Vesicular
transport involves the
transportation offluids with large particles and macromolecules across cellular membranes inside vesicles
(membranous sacs). It is similar to active transport in that it also moves
substances into and out of the cell. Vesicular transport is also used for transcytosis, in which substances are
moved into, across, and out of cells. Vesicular
traffick-ingis a process of movement of substances from anarea (or
membranous organelle) to another. ATP is required for vesicular transport
processes, but another compound (guanosine triphosphate) may also be used. For
transcytosis and endocytosis, protein-coated vesicles allow for movement of
bulk solids, fluids, and most macromolecules.
Endocytosis
Endocytosis andexocytosisboth
use energy from thecell to move substances into or out of the cell with-out
crossing the cell membrane. Relatively large vol-umes of extracellular material
are involved. Energy is required in the form of ATP. In endocytosis, a
secre-tion from the cell membrane moves particles too large to enter the cell
by other processes within a vesicle of the cell. Endosomes are endocytic vesicles. The three forms of endocytosis
are phagocytosis, pinocytosis, and receptor-mediated endocytosis.
Phagocytosis
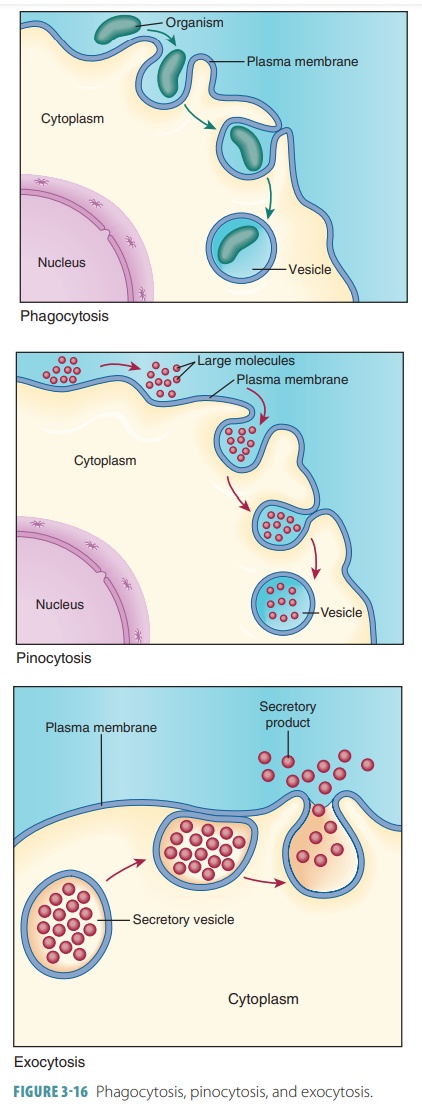
Phagocytosis (“cell eating”) involves cells takingin solids instead of
liquids (FIGURE 3-16). Receptor- mediated endocytosis involves the movement of
specific kinds of particles into the cell, with protein molecules extending
through part of the cell mem-brane to the outer surface. This process is triggered
when a particle binds to receptors on the cell’s
sur-face. Pseudopods (cytoplasmic
extensions) form and flow around the particle. A phagosome (endocytotic vesicle) is formed, which usually fuses
with a lysosome as the contents are digested. Exocytosis is then used to eject
any indigestible contents from the cell. The pri-mary cells used for
phagocytosis are the macrophages and certain white blood cells, commonly
referred to as phagocytes. These
cells ingest and dispose of bacteria,other foreign substances, and dead tissue
cells. Their disposal is important because dead cell remnants can trigger
inflammation or stimulate an unwanted immune response. Phagocytes usually move
via amoe-boid motion, with their
cytoplasm flowing into tem-porary extensions that allow them to propel forward.
Pinocytosis
Pinocytosis (“cell drinking”) involves cells taking insmall liquid
droplets from the surrounding cell envi-ronment with a small indentation of the
cell membrane(FIGURE 3-16). It is also known
as fluid-phase endo-cytosis.
Infolding plasma membrane surrounds extra-cellular fluid that contains
dissolved molecules. This small droplet enters the cell, fusing with an
endosome. It is a routine activity of most cells, which differs from
phagocytosis. Therefore, they can sample the extracel-lular fluid, which is an
important function of cells that absorb nutrients such as those in the
intestines. The parts of the plasma membrane that are removed during the internalization
of the membranous sacs are recycled back via exocytosis. Therefore, the plasma
membrane’s surface area can remain very constant. The endosomes that are formed
by pinocytosis are called pinosomes.
Receptor-Mediated Endocytosis
Receptor-mediated endocytosis is the
primary mech-anism for specific endocytosis and transcytosis of most
macromolecules. Cells use it to focus just on material present in tiny amounts
in the extracel-lular fluid. Plasma membrane proteins that bind specific
substances are used. The receptors and their attached molecules are
internalized in a pit coated with a bristled protein (clathrin). Then,
pinocytosis or phagocytosis occurs. Most receptor molecules are glycoproteins,
each binding to specific targets or ligands.
Receptors bound to ligands usually clustertogether. Endosomes produced when
groove or pock-ets form and move to one cellular area, and then pinch off, are
called coated vesicles due to their surround-ing protein-fiber network. The coating is
required to endosome formation and movement.
In the cell, the coated vesicles
fuse with primary lysosomes containing digestive enzymes to create secondary
lysosomes. Lysosomal enzymes free the ligands from the receptors. The ligands
enter the cyto-plasm via active transport or diffusion. The vesicular membrane
detaches and returns to the cell surface. Its receptors can then bind to more
ligands.
Receptor-mediated endocytosis is
used to take up enzymes, insulin and other hormones, iron, and low-density
lipoproteins such as cholesterol. However, this process can be used to enter
cells by cholera toxins, diphtheria, and the influenza virus. For other types
of vesicular transport, coating proteins such as caveolae may be used. These are flask-shaped or tubular pockets of
the plasma membrane. They capture certain mole-cules in coated vesicles and
used forms of transcytosis.
Exocytosis
The opposite process to
endocytosis is exocytosis, in which a substance stored in a vesicle is secreted from the cell
(FIGURE 3-16). In exocytosis, stimulation occurs via binding of a hormone to a
membrane receptor or because of a change in membrane voltage. Exocytosis is
involved in hormone secretion, mucous secretion, neurotransmitter release, and
sometimes waste ejection. A secretory
vesicle forms, enclosing the substance to be removed from the cell.
Usually, this vesicle migrates to and fuses with the plasma mem-brane. It then
ruptures and its contents are spilled outside the cell. Exocytosis uses a
process wherein transmembrane proteins (v-SNAREs) on the vesi-cles recognize
specific plasma membrane proteins (t-SNAREs). They bind with them, causing the
mem-branes to fuse together in a “corkscrew” pattern. Lipid monolayers are
rearranged without mixing together with the transmembrane proteins. Material
added by exocytosis is then removed by endocytosis.
1. Distinguish
between phagocytosis and pinocytosis.
2. Explain
the three active cell mechanisms.
3. Describe
endocytosis.
4. Describe
exocytosis.
5. Describe
vesicular transport.
Related Topics