Cytoplasm (Cytosol, Organelles) Structure
| Home | | Anatomy and Physiology | | Anatomy and Physiology Health Education (APHE) |Chapter: Anatomy and Physiology for Health Professionals: Levels of Organization : Cells
1. What are the major differences between cytosol and extracellular fluid? 2. Identify the differences between RER and SER. 3. What is the role of the mitochondria? 4. Compare ribosomes and lysosomes
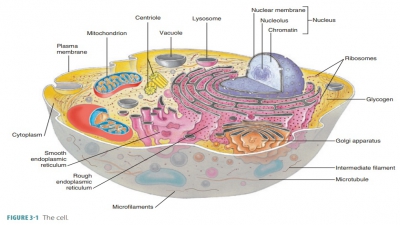
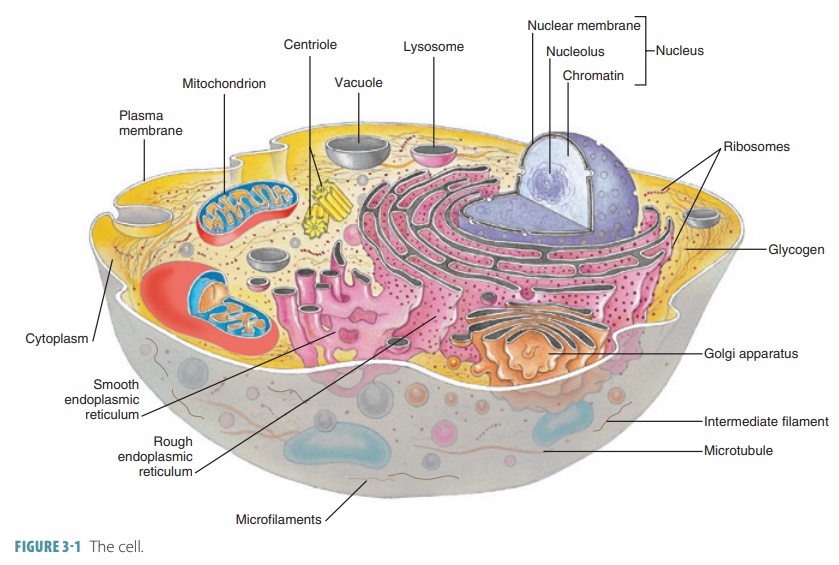
Cytoplasm
Cytoplasm is the substance that contains all the cellular contents
between the cell membrane and the nucleus. It serves as a matrix substance in
which chemical reactions occur. Cytoplasm makes up most of each cell’s volume
and is a gel-like material suspending the cell’s organelles. It usually
appears clear with scattered “specks,” although more powerful magnification
reveals that it contains membranous networks, pro-tein frameworks, and a
cytoskeleton.
Cytoplasm consists of cytosol and
organelles (excluding the nucleus), which are subcellular structures that
perform specific functions.
Cytosol
Cytosol is the fluid portion of
cytoplasm, containing mostly water as well as glucose, amino acids, fattyacids,
ions, lipids, proteins, ATP, and waste products. Cytosol is the site of many
chemical reactions that are required for cells to exist. It is the part of the
cytoplasm that cannot be removed by centrifugation.
The most important differences
between cytosol and extracellular fluid are:
■■ Cytosol contains higher amounts of suspended proteins than does
extracellular fluid. Many of these proteins are enzymes that regulate
meta-bolic operations; others are involved with the var-ious organelles.
■■ Cytosol also contains higher amounts of potas-sium ions than do
extracellular fluid; however, the concentration of sodium ions is much lower in
cytosol than in extracellular fluid.
■■ Cytosol usually contains small amounts of lipids, carbohydrates, and
amino acids.
Organelles
The cytoplasm receives,
processes, and uses nutrients. It contains various types of organelles
(nonmembra-nous or membranous). Organelles perform most of the tasks that keep
the cell alive and functioning normally. Each organelle accomplishes specific
tasks related to cell structure, growth, maintenance, and metabolism. An
organelle’s membrane often allows it to unite with the interactive,
intracellular endomembrane system.
The organelles have specific actions that help the cell to carry out its
activities.
Microtubules
Microtubules are hollow tubes found in most cellsthat are constructed
from a globular protein called tubulin. Microtubules are about 25 nanometers insize, making them
the largest components of the cytoskeleton. They extend out into the cell’s
periph-ery from an area near the nucleus called the centro-some. Microtubules are differently distributed andhave
different amounts, over time. They form because of the aggregation of tubulin
molecules and grow out from their origination at the centrosome. Eventually,
microtubules disassemble into individual molecules of tubulin. The functions of
microtubules are:
■■ Formation of primary cytoskeleton
components:This strengthens cells, makes them more rigid, and anchors the
position of major organelles
■■ Disassembly: When microtubules
disassemble,they help the cell to change shape, which may assist with cell
movement
■■ Movement of vesicles and other
organelles inside the cell: This is related to molecular motors, whichare proteins that bind to structures being
movedas well as to microtubules, moving along their length. Direction of
movement is based on which proteins are involved:
The proteins dynein and kinesin carry materi-als toward the
opposite ends of a microtubule, requiring ATP—these functions are essential to
normal cell function
■■ Formation of the spindle
apparatus: During celldivision, this process distributes duplicated
chro-mosomes to opposite ends of dividing cells
■■ Formation of structural
components of organelles: Including centrioles and cilia
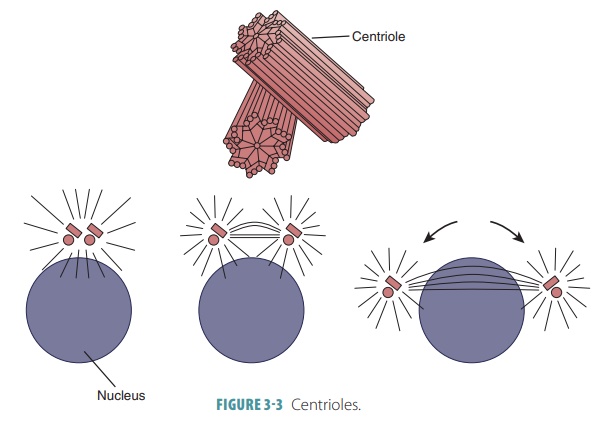
Centrioles
Cell division requires a pair of centrioles, which
are cylindrical structures composed of short microtubules (FIGURE 3-3). During
cell division, the centrioles form the spindle-shaped structure needed for
movement of DNA strands. Cardiac muscle cells, skeletal mus-cle cells, mature
red blood cells, and typical neurons have no centrioles; therefore, these cells
are incapable of dividing.
The centrosome is the cytoplasm surrounding the centrioles. Microtubules of the
cytoskeleton usu-ally begin at the centrosome and radiate through the
cytoplasm. The centrosome is also known as the cellcenter. It consists of nine microtubule triplets that arearranged like a pinwheel. Each microtubule is
con-nected to the next one by nontubulin proteins. The microtubules are
arranged to form a hollow tube. The centrioles also form the bases of cilia and
flagella.
Microfilaments
The smallest of the cytoskeletal
elements, microfilaments are composed of the proteins actin and myosin. Similarly, larger cytoskeletal elements include the intermediate
filaments and microtubules. They are typically found in muscle cells.
Microfila-ments provide cell movement and contraction via interaction with
actin and myosin. This process can also change the shape of the entire cell.
Microfilaments as well as intermediate filaments and microtubules are discussed
in their relation to the cytoskeleton later in this chapter.
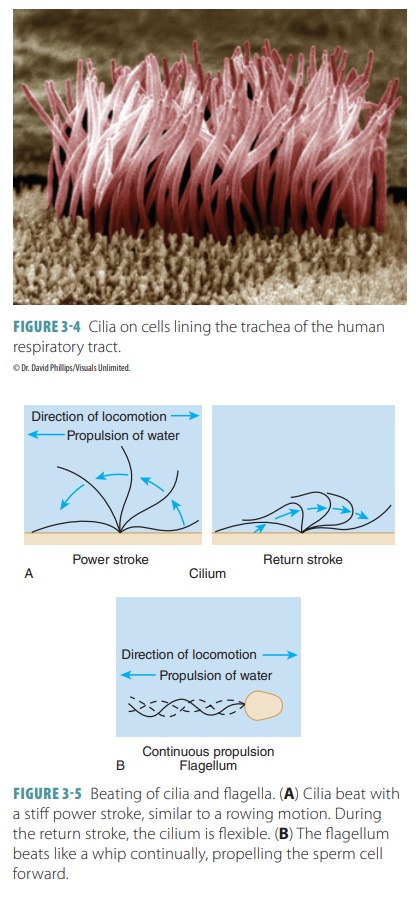
Cilia and Flagella
Cilia, like flagella, extend from certain cell surfaces(FIGURE 3-4). Cilia are short, hair-like structures that move in a
coordinated sweeping motion to propel fluids over the surface of tissues (FIGURE 3-5) . They are found in large numbers on cells lining the
respi-ratory and reproductive tracts. Cilia are formed when centrioles
multiply, lining up beneath the plasma membrane at the cell’s exposed (free)
sur-face. The microtubules emerge from each centriole to form the ciliary
projections. They accomplish this by causing pressure on the plasma membrane.
During this time, the centrioles are referred to as basal bodies. As a cilium moves, it experiences pro-pulsivepower strokes and recovery strokes, which bend and return it to its initial position.
It can repeat these two strokes between 10 and 20 times per sec-ond. When one
cilium bends, it is soon followed bythe
bending of the next cilium, and so forth. This creates a cell surface
“current.”
Flagella are longer than cilia and often exist asonly a single
flagellum. The only example of a flagel-lum is the “tail” of a sperm cell (FIGURE 3-6 ). The key difference between cilia and flagella is that cilia
pro-pel other substances, whereas flagella propel the cells to which they are
attached. There are also nonmotilecilia (primary cilia), which are actually
present just asone single cilium on the surface of most cells in the body. Primary cilia act as antennae, which examine the external
environment for recognizable molecules. They coordinate various intracellular
pathways regu-lating embryonic development and maintain healthy tissues in
later life.
Microvilli
Microvilli are tiny, finger-like extensions of theplasma membrane. They
project from exposed cell surfaces, increasing the plasma membrane surface area
to a large degree. Microvilli are usually found on absorptive cell surfaces,
such as in the kidney tubules and intestines. A core of actin filaments, in
bundles, extends into the terminal web
of the cytoskeleton. In the microvilli, actin appears to have a mechanically
stiffening function.
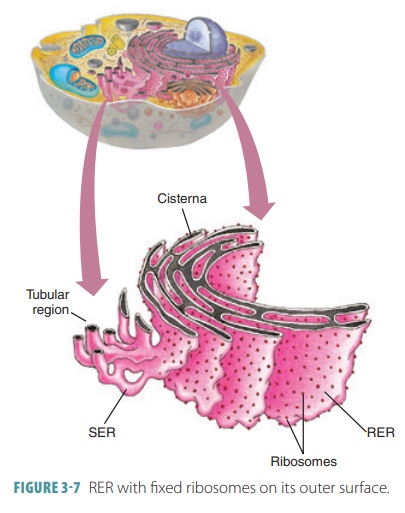
Endoplasmic Reticulum
The endoplasmic
reticulum (ER) is a network of intracellular
membranes connected to the nuclear envelope, which surrounds a cell’s nucleus.
It has interconnected tubules and parallel membranes that enclose fluid-filled cisterns
(cavities). The ER coils and twists through the cytosol and is con-tinuous with
the outer nuclear membrane. Nearly 50% of the cell’s membranes are made up of
the ER. The two types of ER are the smooth
endoplasmicreticulum (SER) and the rough endoplasmic reticulum (RER). The SER does not have ribo-somes on its outer
surface, whereas fixed ribo-somes appear on the RER’s outer surface, giving it“studded” appearance (FIGURE 3-7). Proteins on these ribosomes are threaded into the ER cisterns. The
SER can synthesize phospholipids and cho-lesterol, which are needed for the
cell membrane’s growth and maintenance. It is continuous with the RER,
consisting of a network of looped tubules.
Its enzymes catalyze many
different reactions. These reactions are used for many functions, including
metabolizing lipids, synthesizing steroid-based hormones, detoxification of
drugs and chemicals, breaking down stored glycogen to form free glucose, and
for fat absorption, synthesis, and transport.
Cardiac and skeletal muscle
cells have an elaborate SER (the sarcoplasmic
reticulum) that helps to store and release calcium during muscle
contraction. Overall, most body cells contain very little SER. The RER can
synthesize proteins, and newly made pro-teins are enclosed in vesicles when
they move to the Golgi apparatus for additional processing. In most secretory
cells, liver cells, and antibody-producing plasma cells, the RER is very well
developed. The RER is the cell’s membrane
factory, manufacturing integral proteins and phospholipids that form parts
of cellular membranes. On the external face of the ER membrane, enzymes
required for lipid synthe-sis have active sites. Both free and fixed ribosomes
synthesize proteins via instructions from messen-ger RNA. The amount of ER,
along with the propor-tion of RER to SER, is varied between different cells and
their activities. One example is the pancreatic cells that make digestive
enzymes. They contain an extensive RER, but have a relatively small SER. The
opposite situation exists in the reproductive organ cells that synthesize the
steroid hormones.
Golgi Apparatus
The Golgi
apparatus, also called the Golgi complex, consists of a stack of
several flattened sacs. These “pancake-like” structures are hollow, with
cavities called cisternae inside
them. The flattening of these sacs is caused by a protein complex that pulls
them, when they contain newly synthesized proteins, off the Golgi. Vesicles
from the RER fuse with the con-vex receiving side of the Golgi, which is known
as the cis face. Glycoproteins are
modified inside, with sugargroups being added or deleted and sometimes with
phosphate groups being added. Three or more types of vesicles bud from the
concave trans face of the Golgi
apparatus. Those that contain proteins to be exported pinch off assecretory vesicles (granules). They migrate to the plasma membrane, discharging their
contents from the cell via exocytosis. The enzyme-producing pancreatic cells
are examples of specialized secretory cells that have a prominent Golgi
apparatus. Other vesicles that contain lipids and transmembrane pro-teins are
pinched off by the Golgi apparatus and sent to the plasma membrane or other membranous
organelles. Digestive enzymes are packaged by the Golgi apparatus into
membranous lysosomes that remain in the cell (FIGURE
3-8). The Golgi apparatus has three
main functions: (1) modifying and packag-ing secretions (such as hormones or
enzymes) that are released via exocytosis, (2) packaging special enzymes inside
vesicles for use in the cytosol, and (3) renewing or modifying the cell
membrane.
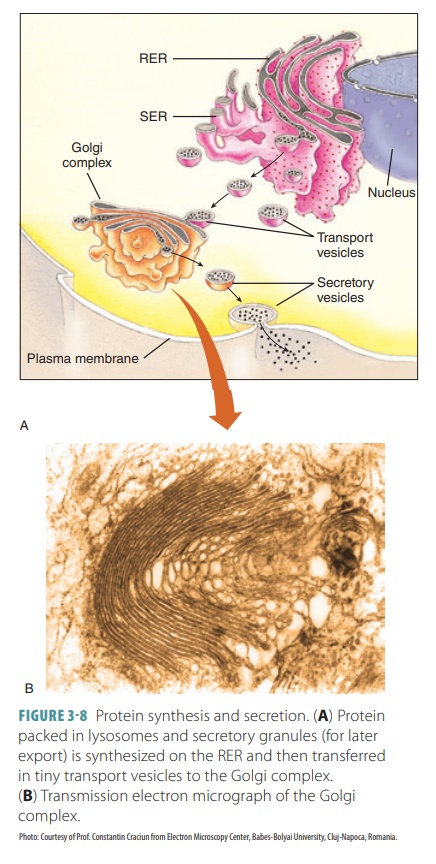
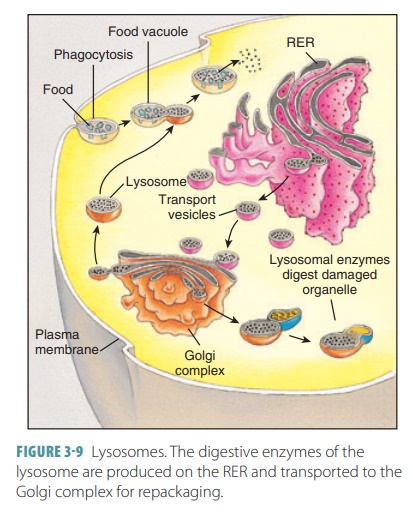
Lysosomes
Lysosomes are tiny spherical sacs that begin asendosomes with inactive
enzymes. They dispose cell wastes, using enzymes to break down nutrients or
foreign particles (such as bacteria). They also destroy older parts of the
cell. This breakdown pro-cess requires the use of powerful enzymes. It often
generates toxic chemicals capable of damaging or killing the cell. Lysosomes
are specialized vesicles that provide an isolated environment for potentially
dangerous chemical reactions (FIGURE 3 -9). They are produced close to the Golgi apparatus and contain
digestive enzymes. In phagocytes, lysosomes are large and plentiful, able to
digest nearly every type of biological molecule. They are most effective in
acidic conditions and are known as acid
hydrolases. The lysosomal membrane contains hydrogen pro-ton pumps. These
ATPases collect hydrogen ions from surrounding cytosol that maintain the acidic
pH of the organelle. The lysosomal membrane also traps dangerous acid
hydrolases, as it allows finaldigestive product to leave for
use by the cell or excretion. Because of lysosomes, sites are provided where
digestion can occur safely inside a cell. The many functions of lysosomes also
include digestion of bacteria, viruses, toxins, and other particles taken in by
endocytosis; performing glycogen breakdown and release and other metabolic
functions; breaking down bone to release calcium into the blood; degrad-ing
organelles that are nonfunctional or “worn out”; and breaking down non-useful
tissues, for example, the uterine lining during menstruation. Although mostly
stable, the lysosomal membrane becomesfragile when the cell is deprived of
oxygen, has too much vitamin A, or is injured. Rupture of lysosomes causes the
cell to digest itself (autolysis), which assists in desirable destruction of cells.
Mitochondria
Mitochondria are thread-like or bean-shaped com-plex membranous
organelles. All cells in the body, with the exception of mature red blood
cells, have between 100 and a few thousand mitochondria (singularly called a mitochondrion). In a living cell, the mitochondria move and change shape on an almost continuous basis.
Mitochondria have double membranes that play a central role in the production
of energy (via ATP). Mitochondria are the “powerhouses” of cells (FIGURE 3-10).
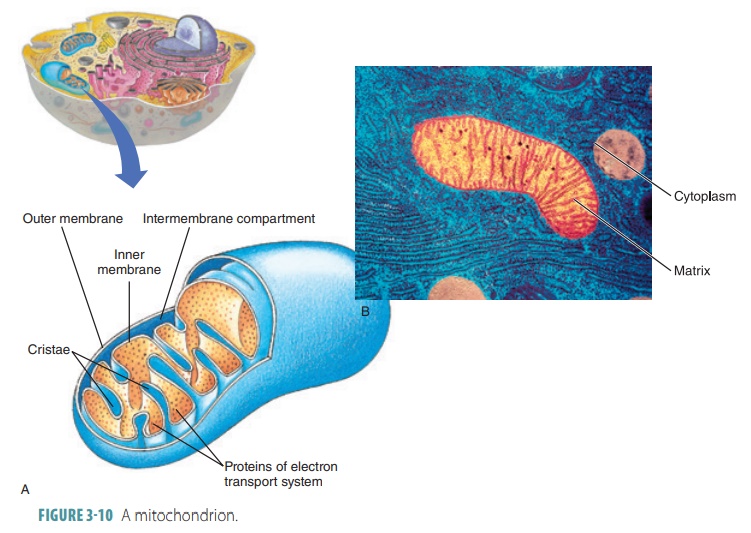
Mitochondria areusually
clustered where most cellular activity occurs. The liver, kidneys, and muscles
have a large number of mitochondria in their cells because they use ATP at a high
rate. A mitochondrion is surrounded by two membranes that are similar in
structure to the plasma membrane. The outer mitochondrial membrane is smooth.
The inner mitochondrial membrane has a series of folds called cristae that protrude into the cen-tral
fluid-filled cavity (the matrix),
which is enclosed by the inner membrane and cristae.
■■ The number of mitochondria in a particular cell varies, based on the
cell’s energy demands.They can migrate through the cytoplasm of a cell and are
able to reproduce themselves. Mito-chondria contain their own DNA, but in a
more primitive form than that found within the cell nucleus. They also contain
their own RNA and ribosomes.
■■Glucose and other food fuel products are broken down by enzymes to
water and carbon diox-ide. Some of these dissolve in the mitochondrial matrix,
whereas others form part of the crista membrane. During oxidization of
metabolites, some released energy is captured and then used to form ATP by
attaching phosphate groups to ade-nosinediphosphate molecules (a process known
as aerobic cellular respiration).
■■Approximately, 37 mitochondrial genes con-trol synthesis of 1% of the
proteins needed for mitochondrial function. The remaining proteins needed for
cellular respiration are encoded by the DNA of the cell nucleus. As the cell
requires more ATP, the mitochondria either halve them-selves (fission) or synthesize more cristae.
This increases their number, and they grow to their former size. Mitochondria
are similar to the purple bacteria phylum. Mitochondrial DNA is also
bacteria-like.
Peroxisomes
Peroxisomes are spherical sacs with enzymes(primarily, oxidases and
catalases) that speed up many biochemical reactions. They are abundant in the
liver and kidney cells, and their diverse actions include synthesis of bile acids, detoxification of hydro-gen peroxide
or alcohol, and breaking down lipids and biochemicals. Oxidases use molecular
oxygen to detoxify alcohol, formaldehyde, and other harmful substances. Most
important, oxidases neutralize freeradicals,
which are highly reactive chemicals. Freeradicals have unpaired electrons that
can ruin the structure of biological molecules. Oxidases convert free radicals
to hydrogen peroxide, which catalyzes quickly into water. Although hydrogen
peroxide and free radicals are normal cellular metabolic byproducts, they can
greatly harm cells if they accumulate in exces-sive numbers. Peroxisomes also
aid in energy metab-olism via the breakdown and synthesis of fatty acids. They
appear similar to small lysosomes that usually form by budding off of the ER
via special processes.
Ribosomes
Ribosomes are small, dark-staining granules that aremade up of
ribosomal RNA and proteins. They are found on the outer membrane of the rough
ER whereprotein synthesis occurs; they
may also be scattered through the cytoplasm. Their functions involve the
formation of proteins, and they are therefore called the “protein factories” of
the cell. They have globular subunits (two per ribosome) that fit together to
form structures that resemble acorns. Protein synthesis is shared by two
different types of ribosomes. Freeribosomes
float freely in cytoplasm, making solubleproteins that function, whereas
other proteins are transported to the mitochondria and certain organ-elles. Membrane-bound ribosomes form the RER and synthesize proteins that will be
incorporated into cell membranes or lysosomes. These proteins may also be
exported out of the cell. Subtypes of ribosomes can change functions. They can
attach to ER membranes as well as detach from them, based on the type of
pro-tein they are making.
Vesicles
Vesicles are also known asvacuoles.These
sacs areformed when a part of a cell membrane folds inward, establishing a
bubble-like structure within the cyto-plasm. Vesicles contain various liquid or
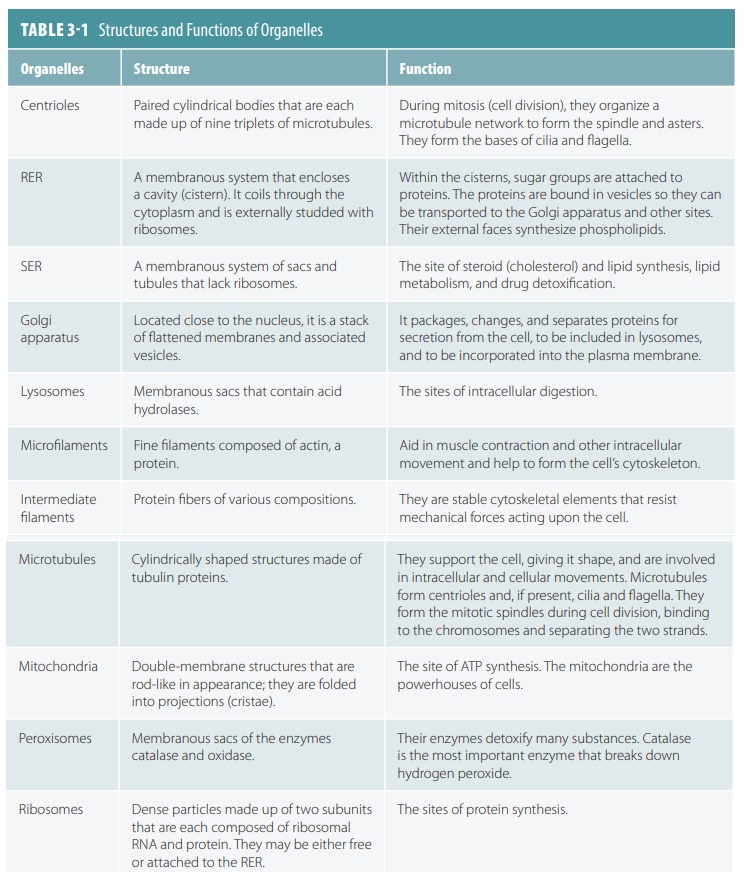
solid materials that formerly existed outside the cell membrane. TABLE 3-1 summarizes the structures and func-tions of organelles.
1. What
are the major differences between cytosol and extracellular fluid?
2. Identify
the differences between RER and SER.
3. What is
the role of the mitochondria?
4. Compare
ribosomes and lysosomes
Related Topics