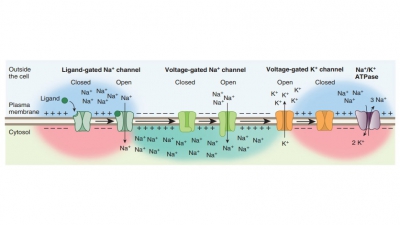
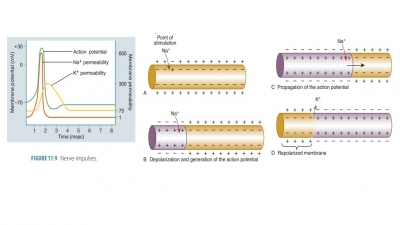
Cells of the Nervous System
| Home | | Anatomy and Physiology | | Anatomy and Physiology Health Education (APHE) |Chapter: Anatomy and Physiology for Health Professionals: Control and Coordination: Neural Tissue
Nerve tissue contains neurons and glial cells (neuroglia).
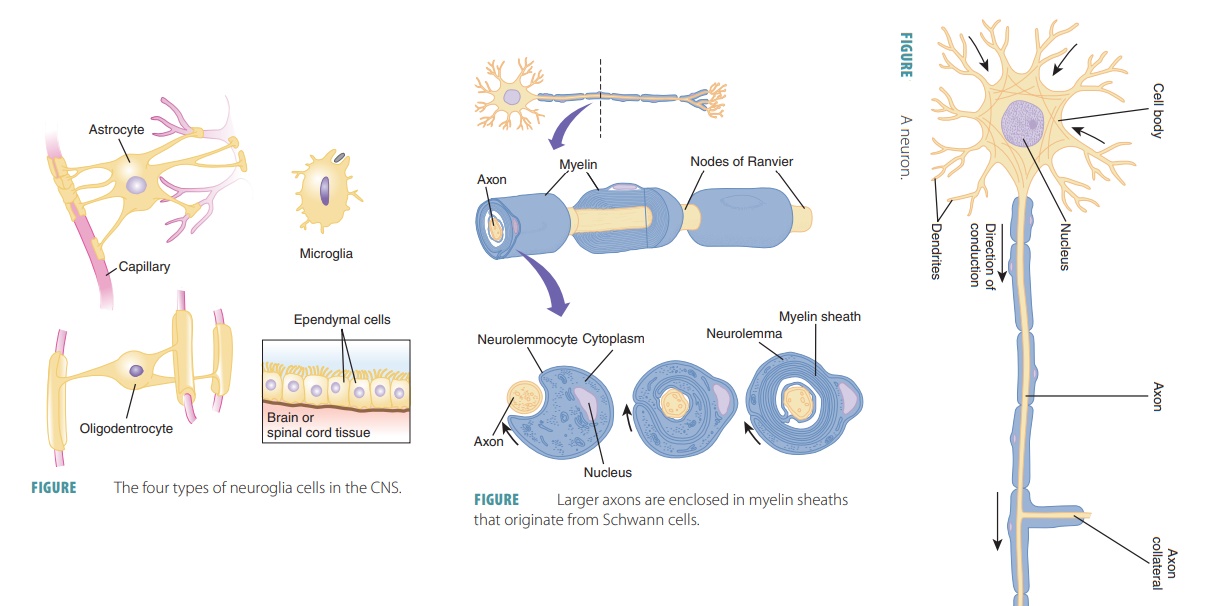
Cells of the
Nervous System
Nerve tissue contains neurons and
glial cells (neuroglia). Neurons are the structural units of the nervous system,
whereas neuroglia support the functions of the neurons. Neuroglia also conduct phagocytosis, fill
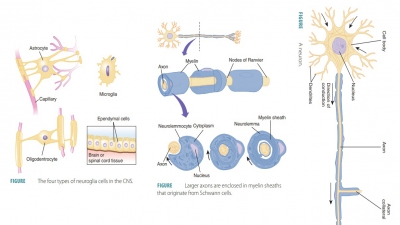
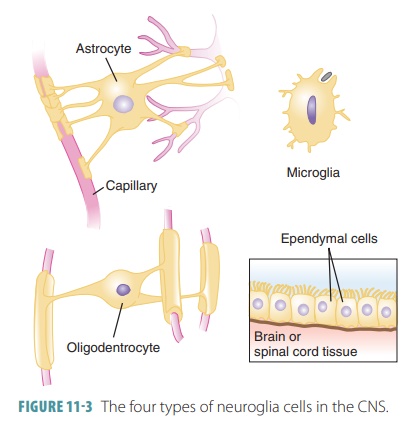
spaces, produce components of myelin, and pro-vide structural frameworks (FIGURE
11-3). There are many more neuroglial
cells than neurons in the body.
Neuroglia
Neuroglia, which exist in both
the CNS and PNS, can divide, whereas most neurons cannot. Most neurog-lia, like
neurons, have branching processes that extend outward and a neutral cell body.
Neuroglia are much smaller than neurons and their nuclei stain dark.
CNS Neuroglia
There are approximately 10 CNS
neuroglia to every one neuron. Neuroglia form approximately half the mass of
the brain. Neuroglia are classified as the fol-lowing types of cells:
Astrocytes
Astrocytes are usually found between neurons
and blood vessels, where they anchor
these compo-nents together. They are star-shaped and delicately branched.
Astrocytes are the most abundant glial cells and also the largest. They have a
variety of func-tions: maintaining the blood–brain barrier, creating the
framework of the CNS, repairing neural tissue damage, and controlling the
interstitial environment as well as the development of neurons. Neural tissue
must be isolated from the body’s general circulation, since its function may be
altered by blood chemicals such as hormones and amino acids. Endothelial cells
that line capillaries control exchanges between blood and interstitial fluid,
creating the blood–brain barrier, to
isolate the CNS from the general circulation.
Many radiating “foot-like” processes grasp neurons and synaptic endings of neurons. They also cover nearby capillaries and aid in exchanges between neurons and capillaries, determining capillary perme-ability. The only breaks in this blanket-like formation occur where other neuroglia contact the capillary walls. Astrocytes secrete chemicals that maintain the amount of permeability of the endothelial cells. Their micro-filaments are significant in number and extend across the cells and their processes. This dense cytoskeleton helps provide the structural framework for the CNS neurons. When damage occurs to neural tissue, it usu-ally does not return to normal, but astrocytes reaching the site of injury can help repair certain structures. This stabilizes the tissue, preventing additional injury.
Astrocytes also control migration
of new neurons as well as the formation of synapses between neu-rons. When a
human is just an embryo, astrocytes are involved in controlling growth and
interconnection of neurons as they develop. Throughout life, the actions of
astrocytes upon the interstitial are many, and include:
■■ Regulating sodium ion, potassium ion, and carbon dioxide concentrations
■■ Creating a rapid transit system so that dissolved gases, ions, and
nutrients may be transported between neurons and capillaries
■■ Controlling volume of blood flow through capillaries
■■ Absorbing and recycling certain neurotransmitters
■■ Releasing chemicals that suppress or enhance communication over
synaptic terminals
Astrocytes also have a vital
function in clean-ing up leaked potassium ions and recycling released
neurotransmitters. Astrocytes also respond to nerve impulses and released
neurotransmitters. They are connected by gap junctions and signal each other
via calcium intake as well as release of extracellular chem-ical messengers.
They create slow intracellular calcium waves and influence neuronal
functioning.
Ependymal Cells
Ependymal
cells line the central
brain and spinal cord cavities, forming a permeable barrier or ependyma between the cerebrospinal
fluid in these cavities
and the tissue fluid around CNS cells. Ependymal cells may range from columnar to squamous and often have cilia. The beating of
these cilia aids in circulation of cerebrospinal fluid, which protects the
brain and spinal cord.
Microglial Cells
Microglial
cells are found throughout
the CNS, and are also called microglia.
They are the least numer-ous and smallest type of neuroglia in the CNS. They
phagocytize bacterial cells and cellular debris. Their phagocytic actions occur
after they transform into specialized macrophages. Microglial cells are oval-shaped
and have lengthy thorn-like processes with many fine branches. These processes
touch neurons that are nearby to monitor their health. When neuro-nal injury or
abnormality is sensed, the microglial cells move toward them. The phagocytic
roles of microglia cells are vital because immune system cells only have
limited CNS access. Microglia appear early in embry-onic development. They
originate from mesodermal stem cells similar to the stem cells that produce
mac-rophages and monocytes. Microglial cells move into the CNS as it forms and
remain there to begin the phagocytic actions.
Oligodendrocytes
Oligodendrocytes
are found aligned along thick nerve fibers and have
smaller cell bodies than astro-cytes. They provide insulating layers of myelin
(the myelin sheath) around axons
within the brain and spinal cord,
insulating the axons from extracellu-lar fluid. They are also branched, but
with fewer processes than astrocytes. These processes are thin cytoplasmic
extensions, mostly contacting exposed surfaces of neurons. The functions of
their processes that end at neuron cell bodies are not understood, but the
functions of processes that end on axon sur-faces are better known.
Near the tip of the
oligodendrocyte processes, the plasma membrane is expanded, forming a large
pad, while the cytoplasm in the area is very thin. Resem-bling a flat pancake,
the structure winds around the axolemma to form concentric layers of the plasma
membrane. This myelin wrap provides electrical insulation and speeds up action
potentials traveling along axons. Large numbers of oligodendrocytes help form
the myelin sheath along the axons, which are then referred to as myelinated. Every oligodendrocyte
myelinates segments made up of several axons. These larger axonal areas,
wrapped in myelin, are called internodes.
Small gaps of only a few micrometers sep-arate adjacent internodes (the nodes of Ranvier). Axo-nal branches
originate at these nodes.
Myelinated axons are glossy
white, mostly because of lipids within the myelin. Therefore, the areas mostly
made up of myelinated axons are called the white
matter of the CNS. The unmyelinated axons of the CNS are not totally covered by neuroglial processes. These axons commonly occur
where short axons and collaterals synapse with dense neuron cell bodies. The
areas made up of neuron cell bodies, dendrites, and unmyelinated axons are
dusky gray in color. This makes up the gray
matter of the CNS. Overall,
oligodendrocytes connect clusters of axons together, aiding in structural
organization. They also improve neuronal functions by wrapping axons within the
myelin sheath.
PNS Neuroglia
In the PNS, there are two types
of neuroglia: satellite and Schwann cells. Satellite
cells have similar func-tions to the
astrocytes of the CNS. They surround neu-ron cell bodies, resembling satellites
around a planet in outer space. Satellite cells are also called amphicytes. Schwann
cells (neurolemmocytes) are neuroglial cells
in the PNS that form a myelin sheath
around axons. Schwann cells do not touch one another, so there are gaps in the
myelin sheath. They play a part in repairing damaged nerves in the PNS. A
process called Walle-rian generation of
an axon that is distal to an injury site results
in macrophages arriving to clean up debris, yet the Schwann cells themselves do
not degenerate. They proliferate to form a cellular cord along the path of the
original axon. In time, the damaged neuron’s axon grows into the site of the
injury, with the Schwann cells wrapping around the axon. Normal synaptic
contacts may or may not be reestablished. This regeneration is not as common
within the CNS.
Brain capillaries are formed by cells that are much more connected than the cells throughout the rest of the body. Partially due to astrocytes, this “high con-nectivity” forms a blood–brain barrier that protects the brain from many chemical substances; for example, certain antihistamines are kept from entering the brain, preventing drowsiness, a common side effect, from occurring. The actions of the Schwann cells are most similar to those of the oligodendrocytes of the CNS.
Neurons
Nervous tissue consists of masses
of neurons (nerve cells) and is highly cellular. In the CNS, the cells are densely
packed and intertwined. Less than 20% of the CNS is extracellular space.
Neurons are the structural and functional units of the nervous system, and each
neuron has a specialized function. Billions of neurons exist in the nervous
system and can function very well for a person’s entire lifetime if they
receive adequate nutrients. However, they are amitotic, losing their ability to
divide, and therefore cannot be replaced if they are destroyed (in most
circumstances). Neurons that can be replaced include the olfactory epithelium
of the nose and certain regions of the hippocampus in the brain, which is
involved in memory. Neurons are larger than other cells of the nervous system
and highly specialized in their conduction of impulses. Nerve impulses are actually electrochemical changes
transmitted by neurons to other neurons
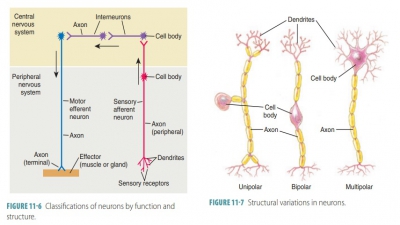
and to cells outside the nervous system. FIGURE
11-4 shows the structure
of a neuron.
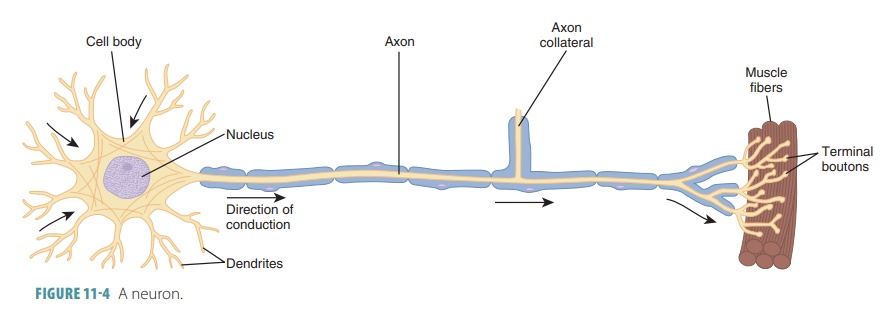
Cell Bodies of Neurons
Each neuron has a rounded cell
body. Although neu-rons are similar in structure, they vary greatly in size and
shape. They all have a cell body or soma, dendrites, and an axon. The cell body, which ranges between 5 and 140
μm in diameter, is made up of a cell membrane, a granular cytoplasm or perikaryon, and
organelles (lysosomes, a Golgi apparatus, mitochon-dria, and fine, thread-like neurofibrils). The
neuro-fibrils are bundles of neurofilaments.
The cytoskeleton of the perikaryon contains these, along with neuro-tubules.
The well-developed Golgi apparatus forms either an arc or a circle around the
nucleus. The cell body is where most biosynthesis occurs in the neu-ron.
Therefore, it contains organelles that synthesize chemicals such as proteins.
To maintain cell integrity and shape, microtubules and the neurofibrils form a
structural network.
Throughout the cytoplasm are many
sac-like Nissl bodies, also known as chromatophilic
substance. These bodies are similar to the
rough endoplasmic reticulum of other cells and stain darkly with com-monly used
dyes. Attached ribosomes synthesize protein. The center of the cell body has a
large, round nucleus with a nucleolus surrounded by cytoplasm. In certain
neurons, the cell body may contain pigments such as black melanin, a red pigment that contains iron, or a gold-brown
pigment called lipofuscin. Most
neuron cell bodies are located in the CNS and are pro-tected by the bones of
the vertebral column and skull. Nuclei are clusters of cell bodies in the CNS, whereas ganglia are clusters of cell bodies in the PNS, which lie along peripheral
nerves.
Processes of Neurons
All neuron cell bodies have processes that
extend outward. These extensions are called dendrites and axons. Dendrites, which may be numerous, receive electrochemical
messages, whereas axons send out electrochemical messages. Each neuron usually
has only one axon. Bundles of axons constitute nerves.
Dendrites
Dendrites have multiple branches that act
as the neu-ron’s main receptive surfaces. The dendrites of the motor neurons are tapered, short in length, and have
diffusely branched extensions. They are
the primary receptive (input) regions of
neurons, having a large sur-face area for receiving neuronal signals. In many
parts of the brain, the finer dendrites are extremely special-ized for
information collection. The dendrites convey messages coming toward the cell
body. These messages are usually short-distance graded potentials instead of long-distance action potentials. A graded potential is also called a local potential. It is a change in the trans-membrane potential that is not
able to spread very far from the area
that surrounds the site of stimulation. The CNS contains both neuron cell
bodies and processes, whereas the PNS mostly contains just processes. In the
CNS, bundles of neuron processes are called tracts. In the PNS, these bundles are called nerves.
Axons
Most neurons have a single axon
arising from an elevation (the axonal
hillock) on the cell body. The axonal
hillock is cone-shaped, narrowing to form a slender process that retains the
same diameter for the remainder of its length. The cytoplasm of an axon is
called the axoplasm, which is surrounded by a spe-cialized portion of the plasma membrane
known as the axolemma. In the CNS, the axolemma may be exposed to interstitial fluid or
covered by neuroglial processes. Neuroglial
cells provide insulation, phys-ical
support, and nutrients to the neurons. Some neurons have short axons or may
even lack axons. In others, axons make up almost the entire neuron length. In
the skeletal muscles of the great toe, axons of motor neurons extend up to 4
feet from the lumbar region of the spine. These are the longest cells of the
body, and long axons such as these are called nerve
fibers.
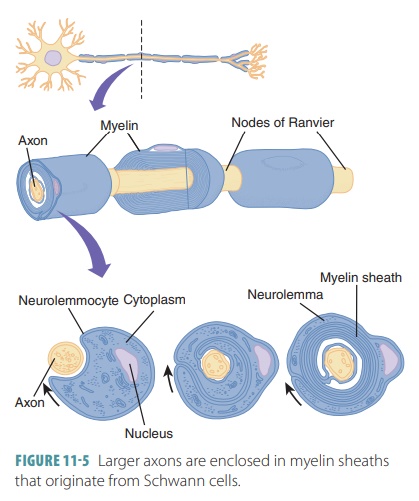
Larger axons are enclosed in myelin sheaths that
originate from Schwann cells ( FIGURE 11-5). These cells are wound tightly around the axons. The areas of the
Schwann cells containing most of the cytoplasm and nuclei are located outside
the myelin sheath, com-prising a neurolemma. Large areas of myelinated axons from oligodendrocytes are known as internodes, which are usually between 1
mm and 2 mm long. The narrow gaps between the myelin sheaths are known as nodes of Ranvier, which
occur at regular inter-vals, approximately 1 mm each, along myelinated axons.
The myelin sheaths are long or have a large diameter. Myelin is a whitish
protein-lipoid. It occurs in segments when it forms the myelin sheath. Myelin
electrically insulates fibers and protects them, increas-ing their transmission
speed of nerve impulses.
Although each neuron has only one axon, some axons have occasional branches known as axon collaterals, which extend at right angles in most cases. Whether an axon is branched or not, it usually has profuse branching at its end (up to approximately 10,000 branches), which are known as terminal branches or telodendria. The distal endings of these terminal branches are knob-like and are called axon terminals, synaptic terminals, synaptic knobs, synaptic boutons, or terminal boutons. When an impulse reaches the axon terminals, it causes neurotransmitter release into the extracellular space. This either excites orinhibits neurons or effector cells that are close to the axon. Axons have the same organelles as dendrites, except for rough endoplasmic reticulum and a Golgi apparatus. Axons decay quickly if they are cut or experience severe damage.
An axon with a myelin sheath is
called myelinated, whereas those without myelin sheaths are called unmyelinated. In the
CNS, dense groups of myelinated axons are white, forming the white matter. The white matter contains
mostly fiber tracts. Those that are unmyelinated, along with neuron cell
bodies, form the gray matter in the
CNS. The gray matter contains mostly nonmyelinated fibers and nerve cell
bodies.
In both the CNS and PNS, axons of
the small-est diameter are nonmyelinated and are covered by long extensions
from adjacent glial cells. Throughout the nervous system, myelinated fibers
conduct nerve impulses more quickly than nonmyelinated fibers. Axons can
regenerate when peripheral nerves become damaged, with the neurolemma playing
an important role. CNS axons are myelinated by oligodendrocytes, which do not
have neurolemmas. When CNS neurons are damaged, they do not usually regenerate.
Col-lections of neural stem cells in the brain can develop new neurons or
neuroglial cells. Neural stem cells are located deep within the brain and near
the brain’s ven-tricles, which contain cerebrospinal fluid.