Synapses
| Home | | Anatomy and Physiology | | Anatomy and Physiology Health Education (APHE) |Chapter: Anatomy and Physiology for Health Professionals: Control and Coordination: Neural Tissue
index: Synaptic Delay, Electrical Synapses, Synaptic Fatigue, Synaptic Activity, Neurotransmitter, Neurotransmitter Chemical Classifications, Neurotransmitter Functional Classifications
Synapses
Nerve
pathways carry nerve impulses.
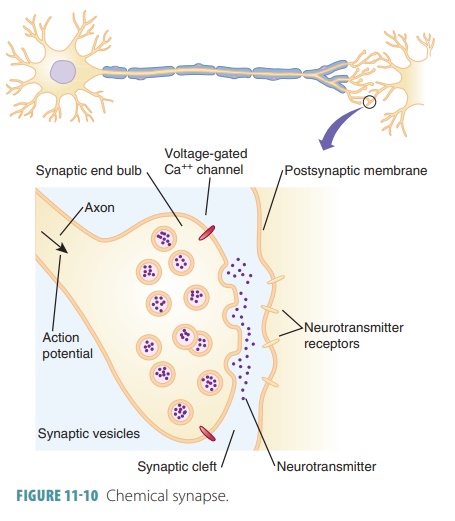
A synapse is a junction between any two communicating neu-rons. The
actual gap between neurons is known as the synaptic cleft. Neurons conduct intracellular commu-nication across these
gaps (FIGURE 11-10). The nervous system requires impulse transmission through
neuron chains that are functionally connected by synapses.
Axodendritic
synapses are those between the axon endings of a
neuron and the dendrites of other neurons. Axosomatic
synapses are those between
axon endings of one neuron and cell bodies (soma) of others. Less common types
of synapses include:
■■ Axoaxonal synapses: Occurring
between axons
■■ Dendrodendritic synapses: Occurring between dendrites
■■ Somatodendritic synapses:
Occurring between cell bodies and dendrites
Neurons may have between 1,000
and 10,000 axon terminals, with synapses being stimulated by an equivalent
number of other neurons. Outside the CNS, postsynaptic cells may be other
neurons or effec-tor cells (gland or muscle cells).
A synapse between a neuron and a muscle cell is known as a neuromuscular junction. Neurons reg-ulate or control the activities of secretory cells at neu-roglandular junctions. Neurons also function in the innervation of many other types of cells, an example of which is fat cells or adipocytes.
A neuron carrying an impulse into a synapse is called a presynaptic neuron. The neuron receiving this impulse is called a postsynaptic neuron. The process of the impulse crossing the synaptic cleft is called synaptic transmission. Most neurons function as both presynaptic and postsynaptic neurons. Syn-aptic transmission occurs in one direction, carried by biochemicals (neurotransmitters).
Chemical Synapses
Chemical
synapses allow the release and
reception of chemical neurotransmitters. Most chemical syn-apses are made
up of two parts: an axon terminal and
a neurotransmitter receptor region.
The axon terminal is a knob-like
structure of the presynaptic neuron that contains many synaptic vesicles, which are very small membrane-bounded sacs holding thousands of
neu-rotransmitter molecules. The neurotransmitter recep-tor region is located
on the cell body or on a dendrite.
The synaptic cleft is a
fluid-filled space that sepa-rates presynaptic and postsynaptic membranes. Each
of these clefts is approximately one-millionth of 1 inch in width. The
electrical current from the presynaptic
membrane dissipates in each synaptic
cleft. There-fore, chemical synapses prevent nerve impulses from being directly
transmitted between neurons. Instead, they are transmitted through chemical
events that are based on release, diffusion, and receptor binding of
neurotransmitter molecules. Neurons, therefore, have unidirectional communication between them.
Complex synaptic terminals exist at each neuro-muscular junction, which
contain parts of the endo-plasmic reticulum, mitochondria, and thousands of
neurotransmitter-filled vesicles. Synaptic terminals reabsorb breakdown
products of neurotransmitters, which are synthesized neurotransmitters from
cell bodies, enzymes, and lysosomes. The movement of these materials is called axoplasmic transport, which may be slow
or fast occurring in both directions. If anterograde, they move from the cell body to the synaptic terminal and
if moving in reverse, they are described as retrograde.
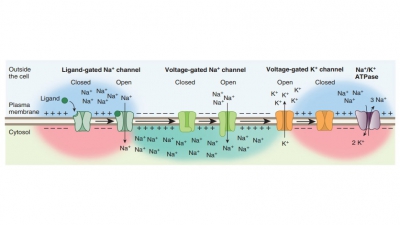
Information is transferred across
chemical syn-apses beginning when an action potential arrives at an axon
terminal. The voltage-gated calcium ion chan-nels then open, allowing calcium
to enter the axon terminal. This entry results in synaptic vesicles releas-ing
neurotransmitter via exocytosis. A single nerve impulse reaching the
presynaptic terminal causes up to 300 vesicles to empty into the synaptic
cleft. A higher impulse frequency causes more synaptic vesicles to fuse and
release their contents. This has a greater effect on postsynaptic cells.
Neurotransmitter diffuses across the synaptic cleft, binding to certain
receptors on the postsynaptic membrane.
This binding opens ion channels to create graded potentials, after which the
neurotransmitter’s effects are terminated.
The events that occur at a
cholinergic synapse, involving the release of ACh, begin with the arrival of an
action potential, which depolarizes the synap-tic terminal. Extracellular
calcium ions enter the syn-aptic terminal. This triggers the exocytosis of ACh,
which then binds to receptors and depolarizes the postsynaptic membrane. ACh
is then removed by acetylcholinesterase
(AchE). This enzyme breaks down ACh via
hydrolysis into acetate and choline. Acetate is then absorbed and
metabolized by the post-synaptic cell or by other tissues and cells. Choline is
actively absorbed by the synaptic terminal so more ACh can be synthesized. This
requires use of acetate that is provided by coenzyme A.
Synaptic Delay
Between the arrival of the action
potential at the syn-aptic terminal and its effect on the postsynaptic
mem-brane, there is a synaptic
delay of between 0.2 msec and 0.5
msec. Most of the synaptic delay reflects the time required for calcium influx
and neurotransmit-ter release, not in the neurotransmitter’s diffusion. The
synaptic cleft is thin and neurotransmitters dif-fuse across it very quickly.
During this time, an action potential is able to travel more than 7 cm (3
inches) along a myelinated axon.
In the CNS, the total synaptic
delay may be lon-ger than the propagation time along axons. Therefore,
reflexes, which are important for survival, utilize only a few synapses,
allowing for rapid, automatic responses to stimuli. The less synapses involved,
the shorter the total synaptic delay, meaning the faster the response. In our
bodies, the fastest reflexes have only one synapse, and a sensory neuron
directly controls a motor neuron.
Electrical Synapses
Electrical
synapses are less common than chemical synapses, consisting of gap junctions similar to those between certain other cells of the
body. Channel pro-teins called connexons
connect cytoplasm from nearby neurons, allowing ions and smaller molecules to
move directly between neurons. These electrically
coupled neurons allow for rapid transmission across electrical synapses.
Communication is unidirectional or bidi-rectional, based on the nature of each
synapse. While synaptic clefts of chemical synapses act as barriers between neurons, electrical synapses allow ions to move directly from
one neuron to another.
This means that all
interconnected neurons have synchronized activities. The adult human brain has
electrical synapses in areas responsible for specific ste-reotypical movements,
such as those that control jerky movements of the eyes. Electrical synapses
also make up the axoaxonal
synapses of the hippocampus, which is
involved in memory and emotions. In embry-onic nervous tissue, electrical
synapses are much more prevalent, allowing exchanging of “cues” that guide
neurons to connect correctly with each other, during early neuronal
development. Over time, chemical synapses replace electrical synapses in
certain areas, becoming the large majority of all synapses in the body.
Synaptic Fatigue
Synaptic
fatigue occurs when intensive
stimulation causes the resynthesis and transport mechanisms of ACh to be
unable to keep pace with demands for this neurotransmitter. Before synaptic
fatigue occurs, mol-ecules of ACh are recycled, and synaptic terminals are not
totally dependent on ACh that is synthesized in cell bodies, and delivered via
axoplasmic transport. After synaptic fatigue, synapses weaken until ACh has
been replenished.
Synaptic Activity
Graded potentials that develop in
the postsynap-tic membrane in response to a neurotransmitter are known as postsynaptic potentials. An excitatory postsynaptic potential (EPSP) is caused by a neu-rotransmitter arriving at the
postsynaptic membrane, due to the opening of the chemically gated membrane
channels, leading to depolarization of the plasma membrane. An inhibitory postsynaptic potential (IPSP) is a graded hyperpolarization of the postsyn-aptic membrane.
It may result from the opening of chemically gated potassium channels, during
which time the neuron is described as inhibited. This is due to a greater
than normal depolarizing stimulus required to bring the membrane potential to
threshold.
Individual EPSPs have tiny
effects on transmem-brane potential, but when they combine through the process
of summation, their effects become inte-grated. There are two types of summation: temporal summation and spatial
summation. Temporal summation occurs when stimuli are added
in rapid succession at just one
synapse that is repeatedly active. Although a typical EPSP lasts only 20 msec,
with max-imum stimulation, an action potential can reach the synaptic terminal
every millisecond. When a second EPSP arrives before the effects of the first
EPSP have disappeared, the effects combine. The degree of depo-larization
continually increases.
Spatial summation involves simultaneous stimuli that are applied at different
locations, cumulatively affecting the transmembrane potential. Multiple
syn-apses are active simultaneously. Each synapse moves sodium ions across the
postsynaptic membrane, to produce a graded potential that has localized
effects. Cumulative effects occur on the initial segment.
Neurotransmitters
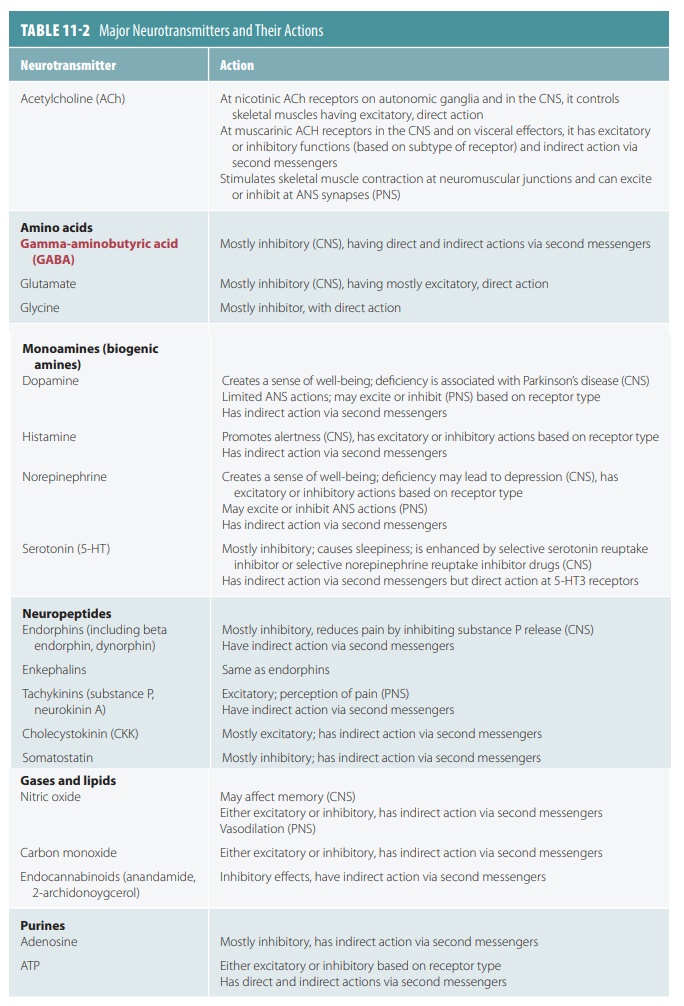
There are about 50 types of neurotransmitters, which are classified chemically and functionally. Neurons may release one or more types. The actions of neurotransmit-ters include effects on sleeping, anger, thinking, hunger, movement, memory, and many other functions. Syn-aptic transmission is commonly affected by either the enhancing or inhibiting effects of neurotransmitters, their destruction, or the blocking of receptor binding. Anything that reduces neurotransmitter activity may slow the brain’s ability to communicate with the rest of the body. Usually, neurotransmitters are released at var-ious stimulation frequencies. This helps to create more ordered synaptic transmission. Simultaneous release of two neurotransmitters from the same vesicle does still occur. Therefore, a cell can have multiple effects on its target. A summary of neurotransmitters and their actions is shown in TABLE 11-2.
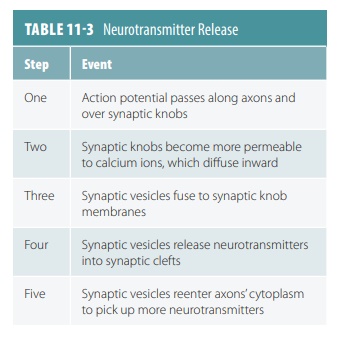
The membrane of a synaptic knob
has increased permeability to calcium ions when an action potential reaches
it. Calcium ions diffuse inward, and in some synaptic vesicles respond by
releasing their contents into the synaptic cleft. Eventually, these vesicles
sepa-rate from the membrane and reenter the cytoplasm to pick up more
neurotransmitters. Neurotransmitters (such as ACh) are decomposed by specific
enzymes (such as AchE) . Others are transported back into the synaptic knobs
that released them in a process known as reuptake. They may also be
transported to neurons or neuroglial cells that are close to them. These
actions prevent neurotransmitters from continually stimulating postsynaptic
neurons (TABLE 11-3).
Neurotransmitter Chemical Classifications
ACh was the first identified neurotransmitter and is released at neuromuscular junctions. The neuromus-cular junctions that release ACh are also known as cholinergic synapses. ACh is synthesized from acetic acid as acetyl coenzyme A and choline via the enzyme cho-line aceyltransferase. It is then transported to synaptic vesicles to be released at a later time. It is then degraded to acetic acid and choline, via the enzyme AchE. Biogenic amines include the catecholamines and indolamines. The catecholamines include dopamine, epinephrine, and norepinephrine. The indolamines include histamine and serotonin.
Amino acids that have identified roles as
neu-rotransmitters include aspartate, glutamate, GABA, and glycine. Peptides or neuropeptides are
basically amino acid chains, and include a mediator of pain signals called substance P and
substances that reduce pain perception while under stress. These substances
include the endorphins and enkephalins. Examples of endorphins include
beta endorphin and dynorphin. Neuropeptides called gut–brain
peptides are found throughout
the gas-trointestinal tract, produced by non-neural body tissues. These peptides include CCK
and somatostatin.
Purines are chemicals that contain nitrogen, which are actually breakdown products of nucleic acids, for example, adenine and guanine. They also include ATP and a part of ATP known as adenosine. Gases and lipids with neurotransmitter actions include gas-otransmitters and endocannabinoids. Examples of gasotransmitters include nitric oxide (NO), carbon monoxide, and hydrogen sulfide. The endocannabi noids are natural neurotransmitters that act at the same receptors as tetrahydrocannabinol, an active ingredient in marijuana. They are lipid soluble and released as needed instead of being stored in vesicles.
Neurotransmitter Functional Classifications
Functional classifications of
neurotransmitters are based on their excitatory or inhibitory actions.
Excit-atory neurotransmitters cause depolarization, whereas inhibitory
neurotransmitters cause hyperpolarization. They are also classified based on
direct versus indirect actions. Those that act directly bind to and open ion
channels. Those that act indirectly cause wider, longer lasting effects via acting
through intracellular second messengers (usually G protein pathways). A
chemical messenger released by a neuron that affects the strength of synaptic
transmission is called a neuromodulator. Most neuromodulators are neuropeptides,
which are small peptide chains that are synthesized and released by synaptic
terminals. They usually act by binding to receptors in the presynaptic or
postsynaptic membranes, activating cytoplasmic enzymes. Opioids are neuromodulators that bind to the same group of postsynaptic
receptors as the drugs opium and mor-phine. The four classes of CNS
opioids are endorphins, enkephalins,
endomorphins, and dynorphins.
Overall, neuromodulators have long-term effects that usually appear slowly and trigger responses that have many steps and various involved compounds. They affect the presynaptic membrane and the postsyn-aptic membrane or both. Neuromodulators also may be released alone or with a neurotransmitter. Many neu-rotransmitters and neuromodulators bind to receptors in the plasma membrane, but require a G protein to link between first and second messengers. As a result, an enzyme called adenylate cyclase may be activated. It converts ATP to cyclic adenosine monophosphate (cAMP) at the inner surface of the plasma membrane. cAMP is a second messenger that can open membrane channels and activate intracellular enzymes or both.
Neuronal Pools
Throughout the CNS, the
interneurons are organized into a smaller amount of neuronal
pools, which are functional groups of
interconnected neurons. These pools may have excitatory or inhibitory effects
on other pools or peripheral effectors. Neuronal pools may be scattered to
involve only neurons in several brain regions or may be localized to just one
certain region of the brain or spinal cord. Output of neuronal pools may
stimulate or depress activities in the CNS. There are five general patterns of
interaction:
■■ Divergence: The spread of
information from a neuron to several other neurons or from one neuronal pool to
additional neuronal pools. It allows broad distribution of specific input.
■■ Convergence: On a single
postsynaptic neuron, several neurons synapse. This means the postsynaptic
neuron can be affected by several patterns of activity in the presynaptic
neurons. Some motor neurons can be under both conscious and subconscious
control as a result of convergence.
■■ Serial processing:
Information is relayed from one neuron to another or from one neuronal pool to
another. For example, the relay of sensory information from one part of the brain
to another.
■■ Parallel processing: When
several neurons or neuronal pools process the same information at the same
time. This means that divergence must occur first. Parallel processing allows
many responses to occur simultaneously.
■■ Reverberation: This resembles
a positive feedback loop, because collateral axon branches along the circuit
extend back to the source of an impulse and stimulate presynaptic neurons
again. A reverberating circuit functions until it is broken by inhibitory
stimuli or synaptic fatigue and can occur in one or more neuronal pools.