Urea Cycle
| Home | | Biochemistry |Chapter: Biochemistry : Amino Acids: Disposal of Nitrogen
Urea is the major disposal form of amino groups derived from amino acids and accounts for about 90% of the nitrogen-containing components of urine.
UREA CYCLE
Urea 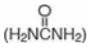 is
the major disposal form of amino groups derived from amino acids and accounts
for about 90% of the nitrogen-containing components of urine. One nitrogen of
the urea molecule is supplied by free ammonia and the other nitrogen by
aspartate. [Note: Glutamate is the immediate precursor of both ammonia (through
oxidative deamination by glutamate dehydrogenase) and aspartate nitrogen
(through transamination of oxaloacetate by AST).] The carbon and oxygen of urea
are derived from CO2 (as HCO3–). Urea is
produced by the liver and then is transported in the blood to the kidneys for
excretion in the urine.
is
the major disposal form of amino groups derived from amino acids and accounts
for about 90% of the nitrogen-containing components of urine. One nitrogen of
the urea molecule is supplied by free ammonia and the other nitrogen by
aspartate. [Note: Glutamate is the immediate precursor of both ammonia (through
oxidative deamination by glutamate dehydrogenase) and aspartate nitrogen
(through transamination of oxaloacetate by AST).] The carbon and oxygen of urea
are derived from CO2 (as HCO3–). Urea is
produced by the liver and then is transported in the blood to the kidneys for
excretion in the urine.
A. Reactions of the cycle
The first two reactions
leading to the synthesis of urea occur in the mitochondrial matrix, whereas the
remaining cycle enzymes are located in the cytosol (Figure 19.14). [Note:
Gluconeogenesis and heme synthesis also involve both the mitochondrial matrix
and the cytosol.]
1. Formation of carbamoyl phosphate: Formation of carbamoyl phosphate
by carbamoyl phosphate synthetase I (CPS I) is driven by cleavage of two
molecules of ATP. Ammonia incorporated into carbamoyl phosphate is provided
primarily by the oxidative deamination of glutamate by mitochondrial glutamate
dehydrogenase (see Figure 19.11). Ultimately, the nitrogen atom derived from
this ammonia becomes one of the nitrogens of urea. CPS I requires
N-acetylglutamate as a positive allosteric activator (see Figure 19.14). [Note:
Carbamoyl phosphate synthetase II participates in the biosynthesis of
pyrimidines. It does not require N-acetylglutamate, uses glutamine as the
nitrogen source, and occurs in the cytosol.]
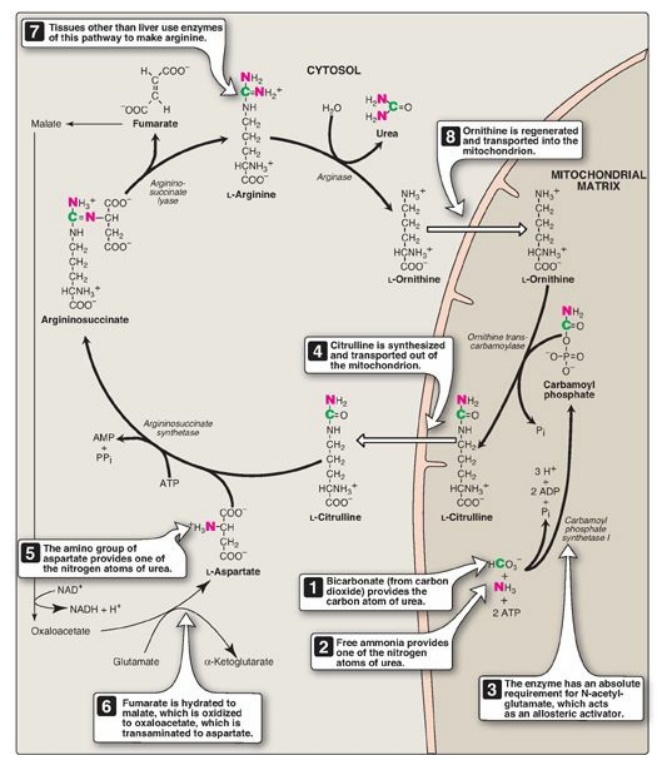
Figure 19.14 Reactions of the
urea cycle. [Note: An antiporter transports citrulline and ornithine across the
inner mitochondrial membrane.] ADP = adenosine diphosphate; AMP = adenosine
monophosphate; PPi = pyrophosphate; Pi = inorganic phosphate; NAD(H) =
nicotinamide adenine dinucleotide
2. Formation of citrulline: The carbamoyl portion of carbamoyl
phosphate is transferred to ornithine by ornithine transcarbam-oylase (OTC) as
the high-energy phosphate is released as inorganic phosphate. The reaction
product, citrulline, is transported to the cytosol. [Note: Ornithine and
citrulline are basic amino acids that participate in the urea cycle, moving
across the inner mitochondrial membrane via a cotransporter. They are not
incorporated into cellular proteins because there are no codons for these amino
acids.] Ornithine is regenerated with each turn of the urea cycle, much in the
same way that oxaloacetate is regenerated by the reactions of the citric acid
cycle.
3. Synthesis of argininosuccinate: Argininosuccinate synthetase combines citrulline with aspartate to form argininosuccinate. The α-amino group of aspartate provides the second nitrogen that is ultimately incorporated into urea. The formation of argininosuccinate is driven by the cleavage of ATP to adenosine monophosphate and pyrophosphate. This is the third and final molecule of ATP consumed in the formation of urea.
4. Cleavage of argininosuccinate: Argininosuccinate is cleaved by
argininosuccinate lyase to yield arginine and fumarate. The arginine formed by
this reaction serves as the immediate precursor of urea. Fumarate produced in
the urea cycle is hydrated to malate, providing a link with several metabolic
pathways. For example, the malate can be transported into the mitochondria via
the malate–aspartate shuttle, reenter the tricarboxylic acid cycle, and get
oxidized to oxaloacetate, which can be used for gluconeogenesis. [Note: Malate
oxidation generates NADH and, subsequently, ATP.] Alternatively, the
oxaloacetate can be converted to aspartate via transamination (see Figure 19.8)
and can enter the urea cycle (see Figure 19.14).
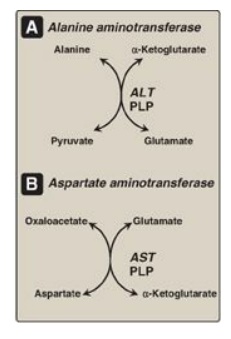
Figure 19.8 Reactions catalyzed during amino acid catabolism. A. Alanine aminotransferase (ALT). B. Aspartate aminotransferase (AST). PLP = pyridoxal phosphate.
5. Cleavage of arginine to ornithine and urea: Arginase hydrolyzes arginine to
ornithine and urea and is virtually exclusive to the liver. Therefore, only the
liver can cleave arginine, thereby synthesizing urea, whereas other tissues,
such as the kidney, can synthesize arginine by these reactions.
6. Fate of urea: Urea diffuses from the liver, and is transported in the blood to the kidneys, where it is filtered and excreted in the urine (see Figure 19.19). A portion of the urea diffuses from the blood into the intestine and is cleaved to CO2 and NH3 by bacterial urease. This ammonia is partly lost in the feces and is partly reabsorbed into the blood. In patients with kidney failure, plasma urea levels are elevated, promoting a greater transfer of urea from blood into the gut. The intestinal action of urease on this urea becomes a clinically important source of ammonia, contributing to the hyperammonemia often seen in these patients. Oral administration of antibiotics reduces the number of intestinal bacteria responsible for this NH3 production.
B. Overall stoichiometry of the urea cycle
Aspartate + NH3
+ HCO3– + 3I ATP + H2O
→
urea + fumarate + 2 I ADP + AMP + 2 Pi + PPi
Because four high-energy phosphate bonds are consumed in the synthesis of each molecule of urea, the synthesis of urea is irreversible, with a large, negative ∆G. One nitrogen of the urea molecule is supplied by free NH3, and the other nitrogen by aspartate. Glutamate is the immediate precursor of both ammonia (through oxidative deamination by glutamate dehydrogenase) and aspartate nitrogen (through transamination of oxaloacetate by AST). In effect, both nitrogen atoms of urea arise from glutamate, which, in turn, gathers nitrogen from other amino acids (Figure 19.15).
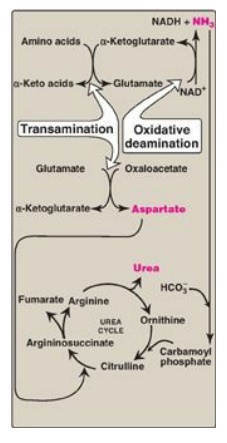
Figure 19.15 Flow of nitrogen from amino acids to urea. Amino groups for urea synthesis are collected in the form of ammonia and aspartate. NAD(H) = nicotinamide adenine dinucleotide.
C. Regulation of the urea cycle
N-Acetylglutamate (NAG)
is an essential activator for CPS I, the rate-limiting step in the urea cycle.
It increases the affinity of CPS I for ATP. NAG is synthesized from acetyl CoA
and glutamate by N-acetylglutamate synthase (Figure 19.16) in a reaction for
which arginine is an activator. The cycle is also regulated by substrate
availability (short-term regulation) and enzyme induction (long term).
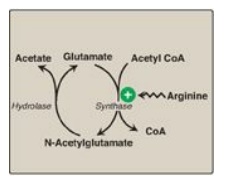
Figure 19.16 Formation and degradation of Nacetylglutamate, an allosteric activator of carbamoyl phosphate synthetase I. CoA = coenzyme A.
Related Topics
